Abstract
A large-scale Ebola viral disease outbreak was identified in Western Africa in early 2014 and the virus has been prevalent in the region since then, resulting in many reported cases and deaths. Global travel, migration, emergence of new infectious agents and their variants increase risks of bioterrorism and pose serious threats to national security and public health. Assays for simultaneous detection of multiple potential bioterrorism agents are of great value, but their current availability is limited. In this study, we report the development of a rapid and specific gold nanoparticle (NP)-based genomic microarray assay (nanomicroarray) for the detection and screening of Ebola, Marburg, and Lassa viruses. Specific capture and intermediate oligonucleotides for Ebola, Marburg, and Lassa viruses were designed and evaluated on the array platform with PCR products derived from these pathogens. The NP-based nanomicroarray assay can detect and distinguish Ebola, Marburg, and Lassa genome sequences in a single assay by displaying different unique patterns on the same platform. The new method described here may be useful for the monitoring of threats from emerging pathogens including new bioterrorism agents and serve as part of emergency preparedness and response to public health.
Keywords
Diagnosis; Filovirus; Hemorrhagic Fevers Viruses (HFVs); Ebola; Marburg; Lassa; Surveillance; Nanomicroarray; Bioterrorism agents
Abbreviations
NP: Nanoparticle; RT-PCR: Reverse Transcription Polymerase Chain Reaction; EVD: Ebola Virus Disease; MVD: Marburg virus disease; HFVs: Hemorrhagic Fevers Viruses; NATs: Nucleic Acid-based Tests; MTA: Materials Transfer Agreement; NGS: Next Generation Sequencing
In 2014, a large-scale Ebola viral disease (EVD) outbreak was identified in Western Africa resulting in over 28,616 reported cases and 11,310 deaths [1,2]. The outbreak began in Guinea in December 2013 and spread to neighboring countries including Liberia, Nigeria, and Sierra Leone. EVD also spread to other countries or continents through the infected travelers and EVD cases have been reported in European countries and the United States [1,3]. According to the WHO report on February 12, 2018, the EVD case fatality rates varied from 25% to 90% in past outbreaks (average fatality rate 50%) [4]. As the symptoms of EVD in the early stage resemble those of common influenza, it is crucial to correctly diagnose the disease using rapid, sensitive and accurate diagnostic techniques to ensure that patients receive timely treatment to prevent exacerbation of the disease, alleviate symptoms and boost immunity for elimination of the virus. The symptoms of EVD start to appear anywhere from 2 to 21 days after exposure to the virus. In May 2018, a new EVD epidemic outbroke in northwestern Congo’s Equateur province and may be posing serious risks for the local population as well as international travelers to the area. A high protective efficacy Ebola vaccine rVSVZEBOV has been developed in clinical trials since the 2014 outbreak [5,6] and distributed in its first phase in May 2018 for EVD emergency preparedness. Among the available EVD tests, Polymerase Chain Reaction (PCR) is a powerful molecular diagnostics tool that can meet the challenges of EVD detection. However, it is less favorable for diagnosing EVD in resource-limited settings due to its costs and requirement for stable working conditions and well-trained professionals.
Self-awareness, preventative measures, and disease monitoring are essential approaches to control the transmission of deadly diseases such as EVD and protect the public health [7]. Following the anthrax attacks in the US in 2001 [8], these measures have been adopted by the U.S. government to manage infectious agents that can be used as biological threats, including smallpox, anthrax, plague, botulism, and the causative agents of Hemorrhagic Fevers Viruses (HFVs). The viruses such as Filoviruses (Ebola and Marburg), Arenaviruses Lassa, Junin, Machupo, Guanarito, and Sabia are listed by the CDC as category A agents [9]. These viruses are widely distributed in nature, can easily infect human beings, and are hard to clinically differentiate from other disease processes [10]. For example, Filoviruses like Ebola and Marburg may exist in some species of fruit bats, infected patients or primates and are transmitted by direct contact with blood, organs, or other bodily fluids of infected hosts [7,11]. The WHO reporter on October 18, 2017 indicated that the case fatality rates of Marburg Virus Disease (MVD) varied from 24% to 88% in past outbreaks (average fatality rate 50%) [12]. It is unfortunate that no proven effective medicines or vaccines exist to help the fight against MVD. The Ebola virus is still active in the body even after the death of the host [13] therefore new infections can occur through contact with tissues or body fluids from a deceased host. Because of their high mortality rates and potential to be used as a bioweapon, all these viruses are listed as bioterrorism agents [7,14]. Many viral and bacterial illnesses can have similar symptoms to viral hemorrhagic fever, therefore, correct identification of the infection and co-infection caused by these different types of pathogens is a great challenge during disease outbreaks.
The standard protocol for screening, diagnosis, subtyping, and discrimination of multiplex co-infections in clinics requires the use of different sets of primers and multiple reactions. Since the last Ebola virus outbreak, many immunoassays and Nucleic Acid Tests (NATs) were developed for the diagnosis of the virus, such as RealStar® Zaire Ebola virus RT-PCR Kit (Altona Diagnostics, Hamburg, Germany) [15], GeneXpert® Ebola Assay (Cepheid, Sunnyvale, California, USA) [16], FilmArray® BioThreat/Ebola Panels (BioFire, Salt Lake City, Utah, USA) [17] and ReEBOV Antigen Rapid Test (Corgenix, Broomfield, Colorado, USA) [18]. These assays can effectively detect Ebola virus in patient serum, plasma, or whole blood and the RT-PCR assay can detect Ebola virus 1 to 2 days earlier than the antigen capture assay [19]. However, NATs may be ineffective for newly emerging viral subtypes due to mutation or mismatch of PCR primers, thus preventing its application in clinics. A study reported two multiplex PCR & RTPCR enzyme hybridization assays (mPCR-EHA, and mRT-PCREHA) for the simultaneous detection of CDC Category “A” DNA and RNA bioterrorism agents, however, that assay requires bio-enzymatic reactions and a standard thermocycler [20]. To exploit the high specificity of the NATs while bypassing its limitations, we developed a diagnostic platform combining nanotechnology and PCR-free whole genomic nanomicroarray. This technique was previously employed for rapid and multiplex detection of influenza virus infections including influenza A avian H5N1 and influenza B viruses [21]. In this work, we adapted the technique for the multiplex detection of common HFVs including Ebola, Marburg, and Lassa viruses. We demonstrated that this platform allowed rapid and simultaneous detection and identification of different HFV genomes in a single test which can directly detect viral DNA or RNA without the involvement of bioenzyme catalyzed amplification reaction.
The PCR products for Ebola (Zaire, 76), Marburg (Variant Musoke), and Lassa (Josiah strain) viruses were obtained from Dr. Jonathan S. Towner (CDC, Atlanta, GA) through the Materials Transfer Agreement (MTA). PCR was performed in a total volume of 30 µL containing 15 µL of 2X PCR buffer (Extensor Hi-Fidelity Reddy Mix PCR Master Mix, ABgen House, Surrey, UK), 2.5 pmol of forward and 2.5 pmol of reverse primers (see Table 1). Reaction conditions were one cycle at 94°C, 5 min, then, 35 cycles at 94°C, 30 sec 50°C, 40 sec 68°C, 1.0 min and one cycle at 68°C for 7 min. The PCR products were quantified using an Agilent 2100 Bioanalyzer (Agilent technologies, Inc., Wilmington, DE) and sequenced using Next-generation sequencing (NGS) prior to detection using the nanomicroarray assay. The details of the NGS assays and bioinformatics were described in our previous publications [21]. All PCR amplicons were accurately and correctly categorized as Ebola, Marburg, and Lassa viruses in individual or mixed PCR products tested. The sequences for capture and intermediate oligonucleotides were designed and prepared following a previous method [21]. Briefly, by using nucleotide sequences available in the National Center for Biotechnology Information (NCBI), we performed multiple sequence alignments of each Ebola, Marburg, and Lassa viral genome with MEGA 5.3 and Vector NTI Advance TM 11 (Invitrogen, Foster City, CA, USA). The selected nucleotide sequences for each of three viral genomes were further aligned to identify significant conserved regions for detection and differentiation of one to others. Multiple capture and intermediate oligonucleotides (usually four or five, respectively) complementary to the conserved regions across the whole-genome or test segments were designed. The PCR fragment for the Ebola virus genome is located between 11624 and 12683 bp (GenBank No: AF086833) while the PCR fragment of Marburg virus genome is located between 6500 and 7757 bp (GenBank No: NC_001608). The PCR fragment for Lassa virus covers the glycoprotein gene between 499 and 1504 bp (GenBank No: M15076). These critical designs ensure that the target gene to be multiply captured on the nanomicroarray and consequently hybridized with multiple intermediate oligonucleotides detected by the gold nanoparticle probe. The designed capture oligonucleotides were modified with 5’-amino-C6-modifier while a 25-mer poly (A) tail added at the 3’ end of intermediate oligonucleotides during synthesis (Integrated DNA Technologies, IA). Capture oligonucleotides that did not bind to any known sequence of Ebola, Marburg, and Lassa viruses were included as array internal positive controls. The oligonucleotides sequences are listed in table 1. Nanomicroarray was designed in a triple-spot format (Figure 1A) and the capture oligonucleotides were printed on CodeLink® activated slides using an OmniGrid Accent® printer (Genomic Solutions Inc., Marlboroug, MI, USA). Procedural details of the nanomicroarray printing, processing, and the assay were described previously [21].
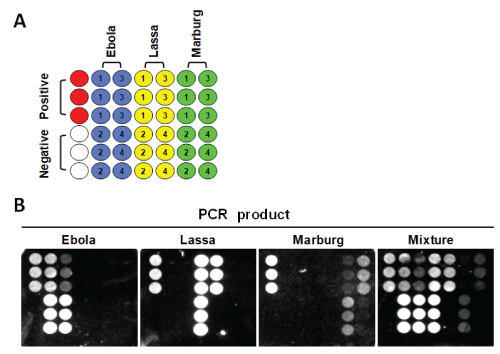
Figure 1: Image from a Verigene IDTM detection system. (A) Nanomicroarray layout with positive control capture (closed red circles), negative control (closed blank circles), Ebola (closed blue circles), Marburg (closed yellow circles), and Lassa (closed green circles) gene captures (there are four different capture oligonucleotides for each gene, and each capture oligonucleotide spotted in triplicate) are indicated. The numbers indicate the different capture sequences from a gene shown in table 1. (B) The capture oligonucleotides detected in each array are labeled as cEbo (for Ebola virus), cLas (for Lassa virus), and cMab (for Marburg virus). Silver staining image for individual or multiple Ebola, Marburg, and Lassa gene’s PCR products are shown.
A nanomicroarray for each gene of Ebola, Marburg, and Lassa virus was designed, printed and evaluated. The nanomicroarray format was fabricated by printing autologous 4 capture oligonucleotides (1 to 4 located from four different regions) in triplicate spots representing a typical virus pattern for Ebola, Marburg, and Lassa as shown in figure 1A. Each subarray contains three gene spots for multiplex assays. The PCR products of three viral genes (1212 bp of Ebola virus, Zaire strain, 76; 1257 bp of Marburg virus, Variant Musoke; and 983 bp of Lassa virus, Josiah strain) were hybridized in the array and the specific signal profiles were observed in the areas printed with corresponding genespecific capture oligonucleotides (Figure 1B). It was observed that PCR amplicons showed a different signal pattern indicating different affinity for each capture oligonucleotide binding to the target template. As shown in figure 1B, cEbo3 shown the lowest affinity to Ebola virus gene; cLas4 showed no affinity to bind to Lassa virus gene but cLas1, 2, and 3 showed strong affinity; the binding affinity for oligonucleotides cMab2 and cMab3 were stronger than the oligonucleotides cMab1 and cMab4 in the Marburg virus gene. The unique fingerprint pattern of the captures for each array represents Ebola, Marburg, or Lassa virus and can correctly identify and differentiate from each other. The different affinity of each capture oligonucleotide may be caused by poor oligonucleotide synthetic or microarray printing process. In the right panel of figure 1B, a mixed PCR product from Ebola, Marburg, and Lassa viruses were tested and a similar image profile for a typical gene was observed in the areas printed with corresponding genespecific capture oligonucleotides. Although there is a different image signal intensity observed in the capture oligonucleotides between the three target genes, however, no cross-hybridization was observed when the single target or multiple targets and multiple intermediate oligonucleotides were incubated in the assay. For laboratory sequence confirmation, the mixture of PCR products was also tested in the NGS assay and each gene was correctly identified (data not shown). We conclude that the current nanomicroarray assay can simultaneously detect and discriminate the three HFVs tested. One benefit of this nanomicroarray assay is that it can randomly and simultaneously detect multiple HFVs in a single subarray from an unknown sample and allow discovery of new emerging strains.
| Oligo ID |
Purpose |
Sequences (5'to 3') |
Locations |
| Ebo_F |
PCR |
CACCAATTGTATTGGACCA |
11624-11642 |
| Ebo_R |
PCR |
AATGTTTGGCAATACTATATTTAAA |
12835-12811 |
| Las_F |
PCR |
TCAGTATGAGGCAATGAGCTGCGA |
499-522 |
| Las_R |
PCR |
AGGCTGTTTGTAGAGTCCACAGGA |
1504-1481 |
| MabF |
PCR |
ACGAATGACACTGGATGCTTCGGT |
6500-6578 |
| MabR |
PCR |
TCCAAGCACTTTGCATGTTCCTCC |
7757-7734 |
| cEbo1 |
capture |
ACTCCCTTAATCCGCAACTACGCAACTGTAAACTCCCGAA |
11684-11723 |
| cEbo2 |
capture |
GGTATCAGGCATCAGTATTCAAAGAAGCGGTTCAAGGGCA |
12215-12254 |
| cEbo3 |
capture |
ATTATGCTCAAAGTACACTGAGAGGAAGGGCCGATTCTTA |
12501-12540 |
| cEbo4 |
capture |
CTTCTCAAGGCACTGTCAGGCAATGGATTCTGTCCTGTTG |
11818-11857 |
| iEbo1 |
intermediate |
AGTTCTTGAGTGATGTACCAGTGGCGACATTGCCCATAGA* |
11762-11801 |
| iEbo2 |
intermediate |
GGTTTGTTCATGATGATTTAATAGACATCTTAGGCTATGG* |
12098-12137 |
| iEbo3 |
intermediate |
ATAGCAGAGATTGAGGATCCAGTTTGTTCTGATTATCCCA* |
12346-12385 |
| iEbo4 |
intermediate |
TGGAGATGACGCCACAACAACTTTGTGAGCTATTTTCCAT* |
12653-12692 |
| cLas1 |
capture |
TTATGAGGATGGCTTGGGGTGGGAGCTACATTGCTCTTGA |
627-666 |
| cLas2 |
capture |
AGACCATCTCCCATCGGTTATCTCGGGCTCCTCTCACAAA |
758-797 |
| cLas3 |
capture |
TACTTGAACGAGACCCACTTTTCTGATGATATTGAACAAC |
1232-1271 |
| cLas4 |
capture |
ACAAGTTTCTATCTTATTAGCATCTTCCTTCACCTAGTCA |
1367-1406 |
| iLas1 |
intermediate |
GAAAGATTAGTGTGCAGTACAACCTGAGTCACAGCTATGC* |
534-573 |
| iLas2 |
intermediate |
AGAGATATTTATATTAGTAGAAGATTGCTAGGCACATTCA* |
803-842 |
| iLas3 |
intermediate |
AAATGACCAACTTATAATGAAGAACCATCTACGGGACATC* |
1090-1129 |
| iLas4 |
intermediate |
CAGCAAGTATTGGTACCTCAACCACACAACTACTGGGAGA* |
1153-1192 |
| cMab1 |
capture |
CTCCGTCCAAAATACCCTCACCACTGCCCACAGCCCGTCC |
6619-6658 |
| cMab2 |
capture |
AGGAACCCTATACAACTTCAGATGCGGTCACTAAGCAAGG |
6766-6805 |
| cMab3 |
capture |
TTCAGCACCCTCTCTGTATCACTACAAAACACCACCAATT |
6990-7029 |
| cMab4 |
capture |
AACACTGCCTACTCTGGAGAAAATGAGAACGATTGTGATG |
7440-7479 |
| iMab1 |
intermediate |
CCAACTGATGCCACCACACTCAACACCACAGACCCAAACA* |
6681-6720 |
| iMab2 |
intermediate |
CCCACAACACCACTGCAATCTCTACTAACAACACCTCCAA* |
6943-6982 |
| iMab3 |
intermediate |
CCAACAGGAAATCTTACCACAGCAAAGAGCACTAACAACA* |
7095-7134 |
| iMab4 |
intermediate |
AGGAGGATGACCTGGCAGCAGGGCTCAGTTGGATACCGTT* |
7504-7543 |
Table 1: Oligonucleotide sequences for capture, intermediate and PCR.
*25-mer poly (A) tail added at 3' end of each intermediate oligonucleotide. Mab: Marburg virus; Las: Lassa virus;
Ebo: Ebola virus; F: Forward; R: Reverse; i: intermediate oligonucleotide; c: capture oligonucleotide
Standard nucleic acid testing provides exquisite sensitivity and specificity, however, prior knowledge of the pathogens’ genomes is necessary for designing pathogen-specific primers and probes, which it is a limitation in identifying novel or genetically modified pathogens. Screening samples for multiple pathogens by real-time PCR is timeconsuming, labor-intensive and requires facilities with high biosafety level, which prevent its applications in common clinics. It was reported that a novel Ebola virus variant with genomic changes developed over time during human-to-human transmission, and the genome variation might affect the efficacy of virus detection and candidate therapeutics [22]. The challenge we face can be greatly improved using the current nanomicroarray platform by multiple capture oligonucleotides targeting wide regions of the target gene and multiple intermediate oligonucleotides by recruiting nanoparticle probes for sensitive detection. As described previously, the nanomicroarray assay can directly detect less than 100 fM for purified PCR fragments and 103 TCID50 units of influenza viral RNA sample without target amplification [23]. We were unable to test genomic viral RNA from Ebola, Marburg, and Lassa virus samples in the current study because of the limitation of materials.
Responding to control emerge bioterrorism agent such as HFVs will require continued investment in the development of diagnostic assays, vaccines, and therapeutic agents. During a bioattack situation, it is possible that a combination of multiple deadly bioterrorism agents may be used. Early diagnosis and differentiation of the specific biological agent as the weapon being used is crucial for bioterrorist attack surveillance to enable implementation of immediate response measures to prevent further spread of the biological agent, to reduce morbidity and mortality, and to minimize economic impact on public health. Because HFVs such as Ebola virus cause common, nonspecific symptoms, large numbers of infected people, particularly those living in the remote regions, may ignore the HFV diagnosis resulting in many unreported cases. To solve this issue, we proposed a diagnostic algorithm for identification of an unknown risk for HFVs using a combination of nanomicroarray and NGS assay platforms. To determine the virus type for a suspected Ebola, Marburg, or Lassa virus infection, viral RNA is extracted from a patient sample and directly analyzed using the nanomicroarray to screen for and determine the specific virus. Once a novel, emerging, or co-infecting virus is found, random reverse transcription PCR is further performed to generate whole-genome amplicons for the positive sample, this amplicon can either be re-tested on the nanomicroarray assay to confirm the initial finding or sent to the central laboratory to perform an NGS assay and data analysis for final sequence confirmation. This algorithm can simultaneously identify, characterize, and discriminate the multiple bioterrorism viral pathogens to initiate immediate response measures to prevent further spread of the biological agent. We can adapt the nanomicroarray assay to a field test by converting the current platform to an inexpensive, rapid and multiplex point-of-care diagnostic platform using microfluidics and biosensor detection technology for viral pathogen detection in clinical settings [24,25]. An optimal assay should include target genes from most HFVs that cover their different types/clades and other bioterrorism agents, and be easily performed by untrained lay users for sample testing in the field in order to serve as an effective preparedness tool to rapidly detect and respond to their exposure. In summary, we have described the development of a nanomicroarray assay to simultaneously detect and distinguish Ebola, Marburg, and Lassa virus infections in a single test. This diagnostic platform could be used to refine current deadly viral surveillance strategies for effective monitoring of these viruses present in circulating populations, early prediction of disease potential, and efficient prevention for improving public health and national security
Acknowledgments
The authors acknowledge Drs Krishnakumar Devadas and Mohan Haleyurgirisetty for their review of the manuscript. "This work was supported by the FDA Medical Countermeasures Initiative (MCMi) funds: #147-09." The findings and conclusions in this article have not been formally disseminated by the Food and Drug Administration and should not be construed to represent any Agency determination or policy.
Article Information
Aritcle Type: SHORT COMMUNICATION
Citation: Zhao J, Liu J, Wang X, Hewlett I (2018) Multiplex Detection of Bioterrorism Agents Ebola, Marburg, and Lassa Viruses using a Nanomicroarray Assay. J Emerg Dis Virol 4(2): dx.doi.org/10.16966/2473-1846.145
Copyright: © 2018 Zhao J, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
Received date: 27 Sep, 2018
Accepted date: 23 Oct, 2018
Published date: 31 Oct, 2018
