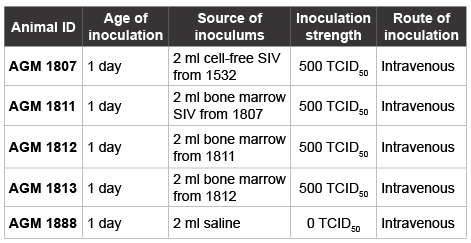
Table 1: TCID50% of SIVagm inoculums used on naïve newborn AGMs

Maria N Kiio1 Valeria N Bosire1 Damian Odoyo1 Elephas Munene1 Joseph Kamau1 Atunga Nyachieo2 Nyamongo Onkoba1*
1Tropical Infectious Disease Department, Institute of Primate Research, Karen-Nairobi, Kenya*Corresponding author: Nyamongo Onkoba, Institute of Primate Research, P.O. Box 24481- 00502, Karen-Nairobi, Kenya, E-mail: bwonkoba@gmail.com
Background: Simian immunodeficiency viruses (SIVs) are diverse group of viruses that naturally infect a wide range of African primates, including African Green Monkeys (AGMs). It has been shown that in vivo passage of SIV in rhesus macaque results in selection of pathogenic virus that causes Simian Acquired Immunodeficiency Syndrome. However, adult AGMs infected with Simian Immunodeficiency Virus (SIV) do not show clinical signs of immunodeficiency.
Objective: Therefore, the present study sought to determine whether pathogenicity occurs in new-born AGMs following serial in vivo passage with SIVagm.
Methodology: One new-born AGM was inoculated with cell-free SIVagm isolated from plasma of a naturally infected AGM and subsequent bone marrow serial transfusions done to other three AGMs at two weeks intervals. Virus isolation and quantification was determined by limiting dilution co-culture method. Virus presence was detected by antigen capture ELISA and polymerase chain reaction. Seroconversion was determined by antibody ELISA and confirmed by Western blot.
Results: Virus isolation and seroconversion confirmed successful SIVagm passage in new-born AGMs. After one year, all the animals did not show clinical symptoms related to SIVagm infection. Bone marrow transfusions from SIVagm infected new-born into naïve new-borns induced persistent infection and anti-SIV antibody response. However, this did not show presence of disease suggesting that AGMs possess inherent resistance that either kill SIVagm or render immune cells of AGMs non-functional.
Conclusion: The results provide valuable information important in investigating the lack of SIV disease pathogenesis in AGMs.
African green monkeys; Simian Immunodeficiency Virus; Serial passage; Pathogenesis
Simian immunodeficiency viruses (SIVs) are a diverse group of viruses that naturally infect a wide range of African primates, including African green monkeys (AGMs) and sooty mangabey monkeys (SMs). Although natural infection is widespread in feral populations of AGMs and SMs the infection generally does not result in immunodeficiency. However, experimental inoculation of Asian macaques results in an immunodeficiency syndrome remarkably similar to AIDS in humans [1].
Over forty African non-human primate species are naturally infected with simian immunodeficiency viruses [2-4]. Among these, African Green Monkeys of Chlorocebus genus are the most numerous, widely geographically spread and most commonly infected with SIV in the wild [2].
In natural hosts, rapid innate immune response to SIV infection during acute infection has been shown to mediate T cell proliferation but lacks capability to regulate SIV replication [5,6]. Innate immunity is activated through induction and up-regulation of interferon responsive genes in both SMs and AGMs [7,8]. Similar phenomenon is observed in a minor subset of HIV-infected individuals who are highly viremic but maintain high CD4+ T cell counts. The genetic profile of T cells isolated from these viremic non-progressors is similar to that of natural hosts; interferon-stimulated gene expression is decreased compared to progressive infection, but similar to chronically infected natural hosts [9].
Various studies on SIVagm infection in AGMs have been undertaken to establish; (i) active viral replication, with set-point viral loads (VLs) similar to or higher than those found in HIV-infected patients [10- 13], (ii) significant depletion of CD4+ T cells during acute infection [11,12,14,15] followed by rapid restoration to near pre-infection levels in the peripheral blood and delayed and incomplete restoration at mucosal sites [10,11,14,16], (iii) maintenance of the balance between Th17 and T regulatory cells, due to preservation of the Th17 cell subset [17], (iv) vigorous but transient inflammatory responses to the virus during acute infection, which are resolved with the transition from acute-to-chronic infection [18], (v) productive infection of short-lived cells [15], (vi) partial control of virus replication by the adaptive immune responses to SIV [19-25] and (vii) CD4+ T cell apoptosis during either acute or chronic infection [26,27]. Therefore, we hypothesize that CD4+ T cell apoptosis together with Th17 cells preservation allows AGMs to avoid enteropathy, breaches in the mucosal barrier and subsequent microbial translocation [14], as well as chronic immune activation and disease progression, while allowing CD4+ T cell recovery in the presence of high viral loads [15].
Furthermore, similar to other natural hosts of SIVs [28], AGMs have a number of adaptations that spare CD4+ T cells from virus-mediated killing in vivo. These features include a lower fraction of CD4+ T cells expressing the CCR5 co-receptor [14,15] and down-regulation of the CD4+ molecule by T helper cells as they enter the memory cell pool [29,30]. Altogether, these adaptations support the concept that the benign course of SIV infection in natural hosts is as a result of co-evolution over millennia [31,32].
However, adequate pathogenesis studies of SIV infection have not been performed to date in wild monkeys due to difficulty in collecting and storing samples on site and the lack of analytical reagents covering extensive SIV diversity [33]. The few pathogenesis studies that have been done support an ancient co-evolutionary relationship by showing that SIVagm are highly adapted to their hosts and SIVagm-infected AGMs generally do not progress to immunodeficiency [15]. It is still unknown why seropositive AGM mothers do not transmit SIVagm virus to their new-born. Thus, the present study utilized naïve new-born AGMs delivered from seronegative mothers to determine whether experimental infection of immunologically naïve new-born AGMs with their natural SIV could lead to pathogenesis.
New-born African green monkeys cercopithecus aethiops delivered from dams seronegative for SIV we sourced from the Institute of Primate Research (IPR) and used in the study. The animals were used within 24-hours of birth and fed daily on commercial starter infant milk formulation (Nestle, Canada) and supplemented with fruits and vegetables. The animals were housed in bio safety containment level 2 facilities at the Institute’s animal holding facility. Study protocols and procedures were approved by nationally recognized Institutional Scientific Ethics and Review Committee and the Animal Care and Use Committee of IPR. The number of animals used in the study was in line with 3Rs principles.
The animals were immobilized using 10 mg/kg body weight Ketamix (Ketamine HCl and xylazine at a ratio of 5:3) during experimental inoculations and sampling. The AGMs were checked daily for general health status, body weight and palpation of the spleen and superficial lymph nodes was undertaken before sampling to check for the effect of the virus on growth and peripheral lymphoid organs.
Newborn AGMs were inoculated intravenously within 24 hours of birth with either 2 ml cell free supernatants from molt-4-clone-8 cells isolate of SIVagm 1532 or bone marrow from previously SIV inoculated AGMs. These newborn AGMs were negative for SIV by ELISA, western blot and virus culture. One newborn (AGM 1807) was inoculated with 500 tissue culture infectious dose 50% of cell free SIVagm 1532 isolated from a naturally infected AGM 1532. The other three newborn AGMs (1811, 1812 and 1813) were inoculated with 105 TCID50 of cell-associated SIVagm virus from previously inoculated newborn. To access the development of immune responses to an unrelated antigen, all the four SIVagm inoculated animals were immunized intramuscularly with 0.2 ml tetanus toxoid in Havlogen at birth and boosted at 16th week throughout the 32nd week of age. The negative control animal AGM1888 was inoculated with 2ml phosphate buffer saline and used as control.
The inoculum used was obtained from plasma of AGM 1532, which was naturally infected with SIV. The virus was grown on molt-4-clone-8 cells for three days and the supernatant propagated on stimulated PBMCs from SIV negative AGM for five days. The strength of SIVagm inoculums used for this study had their TCID50 determined. Briefly, ten-fold serial dilutions of 1ml of plasma in complete RMPI were co-cultured with 106 molt-4-clone-8 cells in 24 well plates in two replicates per dilution (101 - 105 ). The cultures were incubated at 37°C in humidified CO2 incubator and examined twice weekly for the presence of syncytias. Plasma viral load was calculated and expressed as the number of TCID50 per millilitre of plasma. The undiluted culture supernatant was used to inoculate newborn AGMs.
Blood: Heparinised blood was drawn through venipuncture from the femoral vein of each animal on the day of SIVagm administration, 2 and 4 weeks post inoculation and monthly thereafter for twelve months. An aliquot of 200 µl was used for complete blood count. Complete blood counts were performed by a standard automated hematology analyser (Haem Analyzer, Netherlands) for each animal and the hematological parameters compared with standard reference parameters established at IPR for African green monkeys. The rest of the blood was centrifuged for 15 minutes to separate plasma. The plasma was then centrifuged for an additional 5 minutes to remove residual cells. An aliquot of cell-free plasma was used for quantification and isolation of cell-free virus and the remainder stored at -200°C for antibody ELISA and western blot assay. An aliquot of blood was used for isolation of peripheral blood mononuclear cells (PBMCs) by sedimentation gradient separation. A million PBMCs were co-cultured with molt-4-clone-8 cells for virus isolation and quantification. The rest of the PBMCs were stored in liquid nitrogen and at -70°C until further use.
Lymph nodes: One inguinal and/or one auxiliary lymph node was recovered from each animal at pre-inoculation and post-inoculation by transcutaneous biopsy and aseptically teased into single- cell suspensions of lymph node mononuclear cells (LNMCs) in 60 mm diameter tissue culture dishes. The LNMCs were then co-cultured with molt-4-clone-8 cells to recover virus. Briefly, ten-fold serial dilutions of LNMCs in complete RPMI were co-cultured with molt-4-clone-8 cells in 24 well plates in two replicates per dilution (106 -101 ). The cultures were incubated at 37°C in humidified CO2 incubator and examined twice weekly for the presence of syncytia. Culture media were assayed regularly for the presence of SIV major core protein, p27, by antigen capture ELISA. Positive culture results were used to determine cell-associated viral load.
Cell-associated virus load in peripheral blood and cell-free viral load in plasma were determined by limiting dilution culture assays of PBMCs and plasma with molt-4clone-8 cells. Briefly, ten-fold serial dilutions of PBMCs in complete RPMI and two replicates of 101 to 105 PBMCs were co-cultured with 106 molt-4-clone-8 cells in 24-well plates in two replicates per dilution. In addition, 106 PBMCs were co-cultured with 2x106 molt-4-clone-8 cells in T25 flasks. All cultures were incubated at 37°C in humidified CO2 incubator and examined twice weekly for the presence of syncytia. The media was changed every four days and culture supernatants assayed for SIV major core protein p27 by antigen captures ELISA. Molt-4-clone-8 cells were added when the cell concentration appeared to be decreasing. Cultures were maintained for 45 days before termination. Cell-associated virus levels were calculated and expressed as TCID50 per 106 PBMCs. The minimal level of cell associated infectious virus that could be detected was 1 TCID50 per 106 PBMCs. The same coculture procedure was adopted when isolating and quantifying virus in the lymph nodes, spleen, liver, brain, and bone marrow mononuclear cells. Serial ten-fold dilutions of plasma in complete RPMI and two replicates of 101 -105 plasma were co-cultured with 106 molt-4-clone-8 cells in24-well plates. Cultures were maintained as described above. Cell-free viral loads were calculated and expressed as the number of TCID50 per millilitre of plasma or cerebrospinal fluid respectively. The SIVagm were detected by determining the presence of SIV core antigen using ELISA kit in tissue media, plasma or serum. If present, the antigen binds to the antibodycoated micro wells; biotinylated human antibodies that recognize SIV and react with conjugated streptavidin horseradish peroxidase with hydrogen peroxide in the presence of 3’ 3’ 5’ 5’ tetramethylbenzidine substrate. A coloured product was produced and its optical density was measured in a photometer. The culture stopping reagent terminates the reaction and the intensity of the colour developed is directly proportional to the amount of SIV antigen present in the plasma, serum or tissue culture media. The assay was performed according to the protocol provided together with the kit and the absorbency read on a spectrophotometer at 450nm. Samples with absorbency values greater than or equal to the cut off value were considered positive for SIV antigen. Kit manufacturers sum of mean negative control and a pre-determined factor of 0.030 defined the cutoff-value.
DNA extraction: DNA was extracted from cell pellets using the phenol chloroform proteinase-k protocol. A 200 µl of 10% SDS and 5 µl of RNase were added to the cell pellet and incubated for 1 hour at 37°C. A 250 µl of proteinase-k was added and incubated for 5 hours at 55°C. A 200 µl of phenol were added and the mixture span at 3000 rpm for ten minutes. The aqueous layer was transferred into a new falcon tube and DNA extracted twice from this later in 200 µl phenol: chloroform: isoamylalcohol (25:24:1). The aqueous phase was collected into a clean falcon tube and three volumes of absolute ethanol and 10% (v/v) 200 µl of 7.5 M sodium acetate to precipitate the DNA after an overnight incubation at -200°C. The tubes were then span at 3000 rpm for 30 minutes to pellet the precipitated DNA. Excess ethanol was then poured off and the DNA pellet was washed twice in70% ethanol by spinning at high speed for 15 minutes. Ethanol was poured out and the DNA pellet air dried on a blotting paper. The DNA pellet was then suspended in 200 µl of TE buffer and used in PCR assay.
Primer PCR amplification: The reaction was carried out in a DNA thermal cycler to amplify SIVagm provirus. The first round of amplification used primers SIVAGMAGA (5’ AAG TAC CAA ATT AAA CAT TTA ATA TGG GCA GG 3’) and SIVAGMGAGB (5’ CAT TGT CTC TGA TAT GGC CAA ATT TTC CAC A 3’) of the SIVagm gag gene. Briefly, 1 µg of DNA extracted from AGM PBMCs was added to a cocktail round one adjusted to final concentrations of 100 mM Tris-HCl ( pH 8.3); 500 Mm KCl; 1.5 m MgCl2 ; 0.02% gelatin; 200 µl each dATPdGTP and Dttp; 0.5 µl of SIV specific primers and 1 unit of Taq DNA polymerase overlaid with 50 µl mineral oil. The reaction volume was adjusted to 50 µl with triple distilledde-ionised water. After a first denaturation of 4 minutes at 94°C, the reaction was subjected to 32 cycles of amplification. Each cycle consisted of 1 minute denaturation at 94°C, 1 minute annealing at 55°C and 1 minute elongation at 72°C. An aliquot of 5 µl was removed from the primary PCR product and a second round PCR using the internal gag primer pairs, SIVAGMGAGC (5’ CAC CAG GAA AAG AAA GTG AAA GAC ACA GAG GAA GC 3’) and SIVAGMGAGD (5’ GCA TTC TGA ATG AGC AAA GAT TCT GTC ATC CA 3’). The second amplification generated a fragment internal to the first amplified fragment, which allowed an increase in the sensitivity of SIV detection. The final PCR products were detected at 1.5% agarose gel containing 1 µl ethidium bromide against a UV-light background. Positive and negative controls and a DNA molecular weight marker were included in the assay.
Anti-SIV antibodies in plasma were detected by enzyme linkedimmunosorbent assay and confirmed by western blot. Two anti-SIV antibody ELISA assays systems were used to detect anti-SIV antibodies in plasma. One system employed a synthetic SIVmac peptide, as the antigen while the other was a commercial anti-HIV/HIV-2 ELISA kits.
The ELISA protocol used was a modification of a previously described technique. ELISA micro litre plates were coated with a 21 amino acid SIV synthetic peptide derived from the conserved immunodorminant region of the SIVmac transmembrane glycoproteins. The peptide sequence H-NAWACAFRQVCHTTVPWPNAS-OH, was diluted to a final concentration of 0.4 µg/well in bicarbonate buffer. The plates were then sealed, incubated at room temperature for 1 hour then stored at 4°C overnight. The plates were subsequently washed twice and incubated with blocking buffer at 37°C for 1 hour. After washing the plates three times, 100 ml of control and test sera diluted 1:100 was dispensed into the wells in duplicates. The plates were then sealed and incubated at 37°C for 3 hours. After washing, the plates were incubated at 37°C for 2 hours with 100 µl of a 1:2800 diluted goat anti-monkey 1gG conjugate to horseradish peroxide. The plates were washed again and incubated for 30 minutes at room temperature with o-phenylene diamine as the chromogen and hydrogen peroxide as the substrate. The reaction was stopped by addition of 50 µl of diluted sulphuric acid and the optical densities read on a spectrophotometer at 450 nm with reference of 630 nm filter. The cutoff value was defined as the sum of mean negative control and a predetermined factor of 0.100.
The Genelavia commercial kit for indirect enzyme immunoassay for detection of HIV-1 and HIV-2 antibodies in plasma was used (Sanofi Diagnostics Pasteur, Marnes-la-Coquette, France). The plasma samples to be assayed and control sera were diluted at 1:5 with diluents buffer provided with the kit, and 100 µl dispensed into the micro plate wells. The wells were washed and then peroxidase labelled goat-anti-human IgG and IgM antibodies added and incubated in the presence of hydrogen peroxide as the substrate and OPD as the chromogen, after which the excess conjugate fraction had been washed off. The reaction was stopped and the absorbency read on the micro plate ELISA reader. The absorbency measured on a sample allowed the presence or absence of HIV-1 and or HIV-2 cross-reactive antibodies. Samples with absorbency values equal or greater than cut-off value were considered positive. The cut off value was defined as one tenth of mean absorbency of the cut off control serum provided with the kit.
In order to confirm the presence of anti-SIV specific antibodies in the plasma of SIVagm infected newborn AGMs, a new Lav Blot II western blot was performed according to the manufacturer’s instructions. The test is based on the principle of indirect ELISA on a nitrocellulose support containing all the proteins which constitute the HIV-2/SIV virus. Inactivated HIV-2 viral proteins were separated according to their molecular weights by polyacrylamide gel electrophoresis in dissociating and reducing medium and subsequently transferred onto a nitrocellulose membrane sheet. The nitrocellulose membrane sheet was cut into several strips with each strip for running a single test sample/control. These strips were rehydrated and incubated with the test samples and the control serum. If anti-HIV-2 antibodies or cross-reactive antibodies were present, they would bind to the specific virus proteins recognized. After washing off the excess and unbound antibodies, the strips were incubated with alkaline phosphatase labelled anti-human IgG antibodies. The conjugate bound to the anti-HIV-2 or cross-reactive SIV antibodies captured on the slid support. After washing off the unbound and excess conjugate, addition of colour development solution allowed the enzymatic activity of the complex compounds bound to nitrocellulose strips to take place. The appearance of specific coloured bands allowed the presence of antiSIV antibodies in the serum to be detected. The bands were classified as env (gp 140, gp 105 and gp 36) gag (p56,p26, and p16) and pol (p68). According to the manufacturer’s instruction, a test sample is defined positive if it developed at least one band in all the three regions i.e env, gag and pol. SIVagm glycoprotein antibodies cross-reacted with gp 140, gp105 and gp 36 proteins of HIV-2. A sample was defined as negative if no band developed or if the bands appearing could not be classified as env, gag or pol.
Formalin-fixed, paraffin embedded lymph node tissue sections were de-paraffinized and rehydrated with water. Tissues sections were then mounted on microscope slides and dried at 60°C in a dust free oven and stained in haematoxylin and eosin stain. The stained tissues were examined under a microscope and the pathological features in the lymph nodes due to SIVagm infection observed.
The ability of serially passaged SIVagm inoculum to induce viraemia in naïve newborn AGMs was estimated and expressed as TCID50 per 106 PBMCs (Table 1).
Table 1: TCID50% of SIVagm inoculums used on naïve newborn AGMs
At two weeks post-SIVagm inoculation, anti-SIV specific antibodies were detected in plasma of all the SIV-inoculated newborn AGMs. Two anti-SIV antibody peaks were observed at 16th week post inoculation and the at 40th week post SIVagm inoculation. The levels of antibodies were not markedly elevated in plasma of AGM 1811 except a marked peak on the 40th week post SIVagm inoculation. The results showed that AGM 1811 had low antibody response compared to the other three newborns that were subsequently inoculated. A marked low antibody response was observed on the 24th week for all the animals, on the 32nd week for AGM 1812 and AGM1811 and at euthanasia. From the 24th week, AGM 1812 and AGM1813 had low antibody responses compared to AGM 1807 and AGM 1811 (Figure 1).
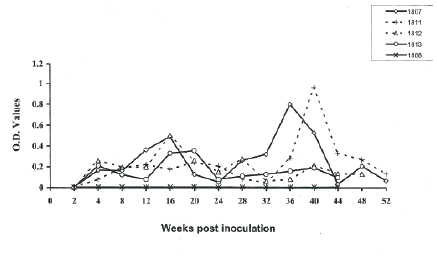
Figure 1: Anti-SIV antibody responses in newborn AGMs inoculated with SIVagm
In all the four SIV-inoculated newborns, hematological parameters examined did not exhibit any nucleotide changes after SIVagm inoculation compared to uninfected controls. All values determined were within the normal ranges observed in uninfected AGMs within the age groups. Except in one animal AGM 1811 where total white blood cell count remained below the normal established range of 4-15 × 103 µl, throughout the study period, no hematological abnormalities were observed in the other SIV-inoculated AGMs (Table 2).
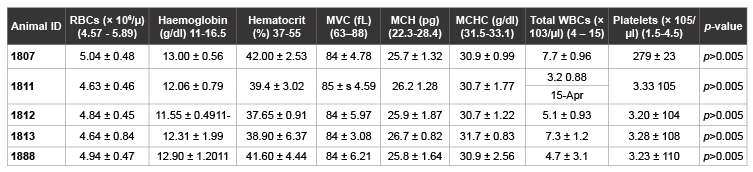
Table 2: Hematologic values in newborn AGMs inoculated with SIVagm
Amplified proviral sequences of SIV gag were detected in AGM PBMC pellets throughout the study. The size of the product amplified by PCR was about 100 base pairs, which corresponded to the molecular weight of gag, the core protein of SIV (Figures 2 and 3).
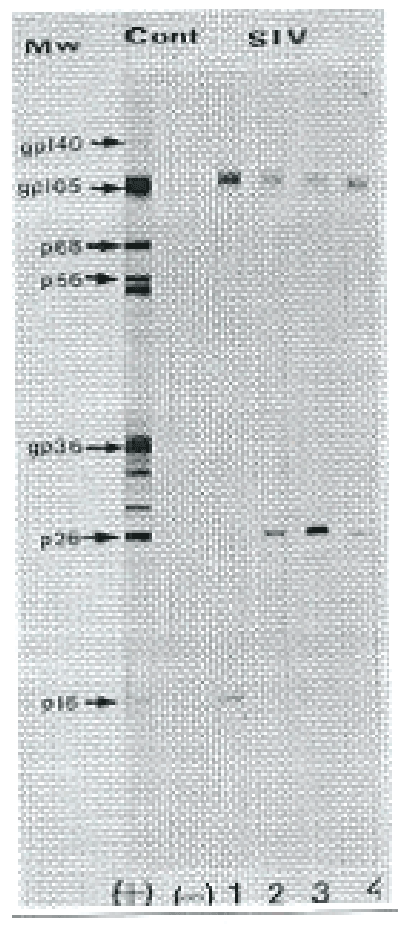
Figure 2: A western blot microphotograph showing presence of SIVagm in AGMs inoculated with SIVagm
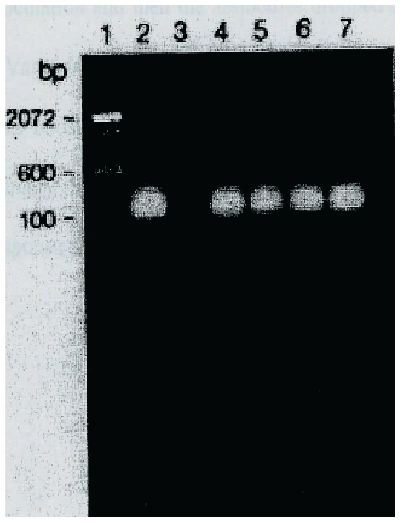
Figure 3: A microphotograph of SIVgag proviral sequence in newborns AGMs inoculated with SIVagm
All four SIVagm newborn AGMs immunized with tetanus toxoid at birth elicited anti-tetanus IgG antibody responses at 2 weeks post SIVagm inoculation and then the antibodies dropped at week16 and lowest at 32nd week post primary SIVagm inoculation and persisted until euthanasia. The primary response was not as strong as the secondary response (Figure 4).
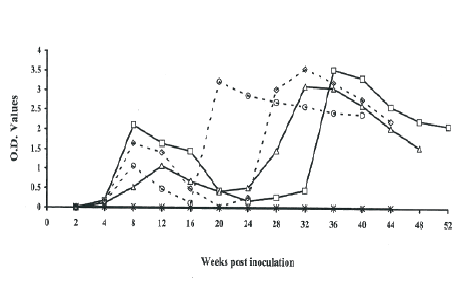
Figure 4: Anti-tetanus antibody response in newborn AGM inoculated with SIVagm
Histopathological examination showed that lymph nodes from the SIV-inoculated newborn AGMs presented with multinucleated giant cells in sinuses and slight swelling. The proviral burden in the lymph nodes was not significantly higher than that observed in PBMCs (Figure 5) or that found in plasma (Figure 6). Lymph nodes showed reactive changes that were mainly affecting the T-cell regions, and encroaching on the germinal centres thereby giving an appearance of loss or reduction in the germinal centres. The changes comprised of a diffuse infiltration of pale staining lympocytic cells and large activated histiocytes. Both T and B-cell types exhibited prominent nucleoli and occasionally mitotic figures could be observed on stained tissues sections. In some instances, progressive transformations of the lymphocytes to immunoblasts were observed. Lymph nodes contained large dysplastic germinal centres of irregular shape that often merged, thereby distorting the structure of the lymphoid tissues. Histologic diagnostics indicated non-suppurative lymphadenitis with para cortical lymphoid hyperplasia at euthanasia. Grading of SIVagm infection of lymph nodes showed AGM 1811 had severe infection, AGM 1812 and AGM 1813 were moderately infected and AGM 1807 lymph nodes presented mild infection. The uninfected control newborn AGM 1888 presented lymph nodes with clearly defined germinal centres, paracortex and cortex. No noticeable alteration of the lymphoid architecture was observed (Figure 7).
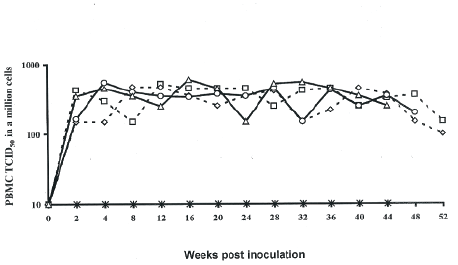
Figure 5: PBMC viral load in newborn AGMs inoculated with SIVagm
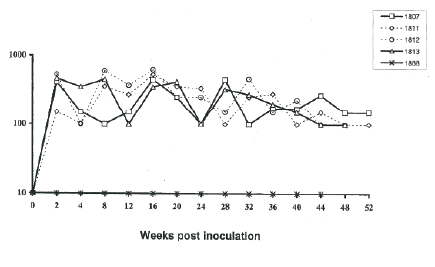
Figure 6: Plasma viral load in newborn AGMs imoculated with SIVagm
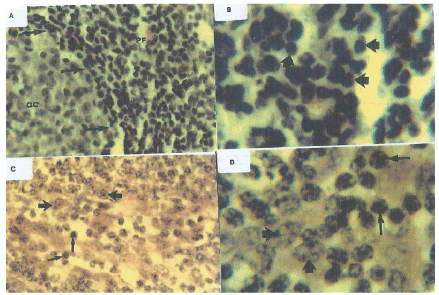
Figure 7: A microphotograph of histopathological features in the lymph nodes
The animals remained clinically healthy throughout the study despite persistent infection, ability to isolate virus and lymphadenopathy. SIVagm inoculated newborn AGMs gained weight throughout the study period, compared to age-matched controls (Figure 8).
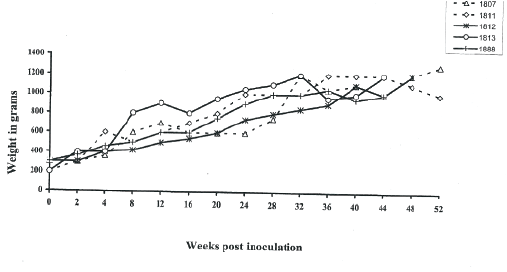
Figure 8: Weight gain in newborn AGMs inoculated with SIVagm
Our findings show that SIVagm was able to induce viraemia in newborn AGMs following serial passage and this was consistent with what has been observed in naturally SIVagm infected adults, HIV-1 infected humans and SIVmac-infected rhesus macaques [34-36]. Viral expression in tissues indicated that the virus was restricted to lymphocytes and macrophages, similar to the distribution observed in SIVmac and SIVagm infection of monkeys [35,37,38]. Earlier studies have indicated that neonatal AGMs have a significantly higher pool of CD4+ T-lymphocytes compared to adult AGMs, it is therefore expected that AGM newborns would be more susceptible to SIVagm infection, which could lead to pathogenesis. However, this was not the case since the viral load in PBMCs and plasma was similar to or lower than those of naturally infected adult AGMs. The low viral load could be attributed to immune complexing of the virus or immunologic suppression of virus infected cells. This is collaborated by what has been observed in another study that showed neonatal viral load to be lower than those observed in adult AGMs [39]. Both adult and newborn AGMs exhibit persistent infection with low viraemia compared to humans infected with HIV-1 [40,41]. This result supports our findings where the newborn AGMs had low viral load with persistent infection during initial stage of infection and indication that SIVagm apathogenic in newborn AGMs similar to adults. We deduce that the SIVagm virus is not controlled to any greater degree than HIV in humans or SIVmac in macaques.
Studies on SIV infection in ‘natural host’ non-human primate species such as SMs and mandrills (Mandrillus sphinx, MND) have revealed that these animals also develop chronic infection with high viral loads [42,43]. However, in stark contrast to pathogenic HIV/SIV infections, natural SIV hosts maintain healthy CD4+ T cell levels and avoid immunodeficiency [44,45]. This peculiarly benign course of infection is thought to reflect a non-pathogenic virus–host equilibrium that has been established over a prolonged period of co-evolution. However, it is unclear whether it’s due to changes in the virus, in the host response or in both. One viral feature that would explain the non-pathogenic infection of natural SIV hosts would be that the virus has evolved a mode of intracellular replication that is not cytopathic for the infected cell. However, recent studies have shown that the in vivo longevity of infected cells in these natural hosts is comparable to that observed in pathogenic infection [46]. Several important differences have also been observed between pathogenic and non-pathogenic infections [47]: natural hosts show lower levels of immune cell activation during chronic infection [48], and lower expression of CCR5 on CD4+ T cells [49].
In our study, we established that there was active viral replication and this is similar to what has been documented in SIVmac-infected macaques and naturally SIV infected adult AGMs. Additionally, we showed that there was disruption lymphoid architecture which is not the case with adult AGMs. Therefore, it does not appear that the reason for SIVagm apathogenicity in adult AGMs is either due to a low viral burden in the lymph nodes or is it due to an intact lymphoid architecture. Hence, neither infection nor subsequent destruction of lymph nodes is the sole reason for immunodeficiency in HIV-infected humans or SIVmacinfected macaques.
There were detectable levels of anti-SIV IgG antibody responses early in the course of infection, which persisted throughout the study. Thus, the newborn AGMs seroconverted to SIVagm infection as evidenced by the sustained antibody responses. This suggests that SIVagm is immunogenic to naïve newborn AGMs similarly to adult SIVagm-infected AGMs [50]. Newborn AGMs were also able make antibodies towards tetanus antigens after SIVagm inoculation. This finding suggests that the SIVagm infection did not result in immunosuppression, nor did it alter the capacity of the immune system to respond in a normal fashion to SIV or tetanus antigens. Age-related host factors that determine the maturation of immunecompetence did not seem to play a role in the rate of disease progression. We were able to isolate the virus from PBMCs, plasma, and other tissues and this shows that the newborn AGMs did mount an effective immune response to eliminate both active and latent viral infection. The lymph nodes showed presence of multinucleated giant cells in sinuses, with reactive changes affecting the T-lymphocyte dependent regions and encroaching on the germinal centres. These observations are in contrast to what had been observed in long-term adult AGMs infected with SIVagm, SIV-infected macaques and HIV-1 infected humans [24]. Our findings show that the destruction of lymphoid architecture and thereby its function is not necessarily due to massive virus infection as observed. Further studies on the lymph nodes of SIVagm-inoculated newborn AGMs need to be explored and thoroughly investigated. In addition, experimentally infected neonatal AGMs should be observed over a long period so as to monitor not only lymphoid changes but also other immunologic parameters. The uninfected controls had no noticeable alteration of the lymphoid architecture and germinal centres and para follicular zones were clearly defined. Therefore, an intact lymphoid architecture does not seem to be the reason for SIVagm apathogenicity in adult AGM, since naïve newborn AGMs inoculated with SIVagm remained healthy despite massive lymphoid destruction. SIV inoculated newborn AGMs gained weight throughout the study period despite persistent infection and slight lymphadenopathy. This is consistent with long-term SIVagm infection of adult AGMs [51,52]. On the contrary, involuntary weight loss has been recognized as a major factor contributing to morbidity and mortality of patients presenting with HIV infection. Rhesus macaques infected with SIVmac lose up to 80% of their body weight for which no known cause has been implicated [40]. Whether this is the reason for SIVagm apathogenicity and hence its lack of morbidity and mortality in SIVagm infected AGMs, remains to be determined.
Transfusion of bone marrow from naturally SIV-infected African green monkeys to naïve newborn AGMs induced persistent infection and elicited an anti-SIV specific IgG antibody response with lymphoid tissue pathology, and weight loss. Furthermore, the infection does not induce immunosuppression of neonatal immune system. However, this is yet no convincing explanation for the apparent resistance of newborn AGMs to disease just as adult AGMs. It seems clear that the lack of a fully developed immune system shortly after birth often cited as the probable cause of higher disease susceptibility in other species does not compromise the AGMs natural resistance to SIVagm induced disease. Therefore, further research is needed to understand SIVagm interaction with its natural host, the AGM.
Download Provisional PDF Here
Article Type: Research Article
Citation: Kiio MN, Bosire VN, Odoyo D, Munene E, Kamau J, et al. (2016) Pathogenesis and Immune Responses in Newborn African Green Monkeys (Cercopithecus aethiops) Inoculated with Simian Immunodeficiency Virus (SIVagm). J Emerg Dis Virol 2(2): doi http://dx.doi.org/10.16966/2473-1846.116
Copyright: © 2016 Kiio MN, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access