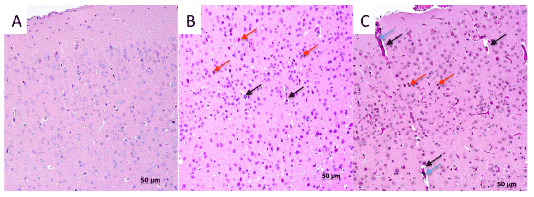
Figure 1: The cerebral cortex of infected mice: A- Control; B – 5 d.p.i.; C – 7 d.p.i. HE (hematoxylin and eosin)-stained sections. Black arrows – swelling; red arrows – necrosis; blue arrows – hemolysis.

Kirill Sharshov1,2,4,* Elena Prokopyeva1,2 Ivan Susloparov2 Wen Wang3 Zhuo Li3 Laixing Li3 Lidia Shestopalova2 Alexander Shestopalov1,2,4
1Research Institute of Experimental and Clinical Medicine, Novosibirsk, Russia*Corresponding author: Kirill Sharshov, 630117, Timakova street/2, Novosibirsk, Russia, E-mail: sharshov@yandex.ru
An outbreak of the Highly Pathogenic Avian Influenza H5N1 (HPAI H5N1) was reported in wild birds in 2010. Three strains (A/black-headed gull/Tyva/8/2010, A/spoonbill/Tyva/1/2010, and A/great crested grebe/Tyva/22/2010) were isolated and studied by the virological and molecularbiological methods. Viruses shown to be highly pathogenic in chicken and mice. We found some disruptions in brain tissues that can be related to the observed neuropathological behavior signs in mice. Close antigenic relationship between the investigated strains and those isolated at the Uvs Nuur Lake in 2009 and in the Russian Far East in 2008 (both clade 2.3.2) were also shown. Phylogenetic analysis of the hemagglutinin gene revealed a close relation to the strains isolated during outbreaks at the same location in 2009, as well as at the Qinghai Lake in 2009, and in Mongolia in 2010; as all of them falling into clade 2.3.2. Early AIV detection in this area can play an important role in terms of outbreak prediction, early warning and isolating new AIV strains when they start spreading from Asia to Russia and Europe. Further investigation of the global distribution of clade 2.3.2 and 2.2 HPAI H5N1 viruses will prove to be invaluable for better understanding of the evolutionary ecology of avian influenza viruses.
Avian influenza; Clade 2.3.2; H5N1 outbreak; Russia; Wild birds
AIV: Avian Influenza Virus; CDC: Center for Disease Control and Prevention; HA: Hemagglutination; HPAI: Highly Pathogenic Avian Influenza; OIE: The World Organisation for Animal Health (Office International des Epizooties); PBS: Phosphate-buffered Saline; PCR (RTPCR): Polymerase chain reaction (Reverse transcription polymerase chain reaction): WHO: World Health Organization
Various outbreaks of the Highly Pathogenic Avian Influenza H5N1 (HPAI H5N1) have been reported at the Uvs Nuur Lake (Tyva Republic, Russian Federation, 50°18’N 92°42’E) on the Russian-Mongolian border (Central Asia) since 2006 [1-3].
Uvs Nuur Lake is a highly saline lake in an endorheic basin-Uvs Nuur Basin in Mongolia with its small part belonging to Russia. It is the largest lake in Mongolia in terms of surface area (3, 350 km²), located at 759 m above the sea level. The water in this lake is highly saline (18.8 ppt), primarily due to the sulphate and sodium ions [4]. Uvs Nuur Lake is an important breeding and stopover area for the different migratory waterfowl. Some wild bird species migrate along the Central Asian Flyway and breed at the Lakes of China, Mongolia and Russia [5], where H5N1 outbreaks occurred.
In June 2006 an outbreak of HPAI H5N1 was reported in wild birds at the Uvs Nuur Lake in Western Siberia, Russia. Virus has affected the following bird species: Great Crested Grebe (Podiceps cristatus), Little Grebe (Tachybaptus ruficollis), Black-headed Gull (Larus ridibundus), Common Pochard (Aythya ferina), and Spoonbill (Platalea leucorodia). One strain, A/duck/Tuva/01/06, isolated during the outbreak was highly pathogenic in chicken and mice and belonged to the Qinghailike group (clade 2.2) [3].
In June 2009, another HPAI H5N1 outbreak was reported in the wild birds in Mongolia (Doitiin Tsagaan Lake and Doroo Lake) and at the Uvs Nuur Lake [2,6]. The bird species involved in this outbreak were the same as in 2006. Phylogenetic analysis of the hemagglutinin gene showed that the detected viruses belonged to clade 2.3.2. A/black-headed gull/Tyva/115/09 and the A/great crested grebe/Tyva/120/09 viruses were clearly distinguishable from the H5N1 viruses previously isolated in this region in 2006, but showed 99% identity with A/whooper swan/ Mongolia/8/2009 and A/whooper swan/Mongolia/2/2009, isolated in Mongolia at the same time [7]. All of the isolated viruses were lethal in both mice and chicken.
Therefore, this study aims to study the continuing outbreaks of H5N1 virus at the Uvs Nuur Lake.
Cloacal swabs and organ samples were tested for influenza viruses by inoculation into the allantoic cavity of 10-day-old embryonating specificpathogen-free chicken eggs according to standard procedures [8]. Each sample underwent at least three passages in chicken eggs, and influenza isolates were identified by both HA assay [8] and real-time RT–PCR («AmpliSens Influenza virus А H5N1-FRT» commercial kit, Russia). All virus isolation attempts were conducted in a biosafety level-3 facility
RNA was extracted from influenza A virus-containing allantoic fluid by using a commercial kit (SV Total RNA Isolation system; Promega, Madison, WI). After reverse transcription, cDNAs were amplified by PCR with PyroStartTM Fast PCR Master mix (Fermentas, Foster City, CA). The identification of HPAI was made using «AmpliSens Influenza virus А H5N1-FRT» commercial kit, Russia. Template DNA was sequenced using BigDye Terminator version 3.1 (Applied Biosystems, Inc., Foster City, CA) using the method described by Hoffman et al. [9]. Samples were analyzed using the Vector NTI 10.0 (Invitrogen, Carlsbad, CA) software package. MEGA 4 (Center for Evolutionary Functional Genomics, The Biodesign Institute, Tempe, AZ) was used to produce phylogenetic trees [10]. For the comparisonHA gene sequences in the GenBank database were used to find the most closely related viruses and they were selected for inclusion in the phylogenetic trees.
The hemagglutination inhibition (HI) test was performed as previously described [8]. Viruses were tested with a hemagglutination inhibition assay with reference antiserum and antiserum against viruses, previously isolated in Russia.
Intravenous pathogenicity of isolates for chickens was determined according to guidelines established by the World Health Organization [8].
Nine-week-old BALB/c mice were lightly anesthetized with ether, inoculated intranasally with 50 µl of successive tenfold dilutions of viruses suspended in PBS, and observed for 20 days. Control mice were inoculated with PBS. Mouse organs (lungs, brains, livers, spleens, kidneys) were collected from each infected group (n=3) according to a pre-designated schedule 1, 3, 5, 7 and 10 days post infection (d.p.i.). Uninfected controls were humanely sacrificed on 10th d.p.i. Sacrification was performed using cervical dislocation. All procedures were performed in accordance with study protocol approved by Bioethical Board.
Affected mice were sacrificed and tissue samples were taken for morphological investigation as described by Potapova et al. [11].
We hypothesized that the bodies of water like the Qinghai Lake and the Uvs Nuur Lake may play an important role in the circulation of the avian influenza so we suggested enhancing the existing surveillance program in this area and continued to study the new outbreaks thoroughly [7]. We conducted continuous surveillance in the reported area including breeding period and period of migration. We looked for bird carcasses and sampled live and dead birds.
Our hypothesis was confirmed in June 2010 when an outbreak of HPAI H5N1 virus was again reported at Uvs Nuur Lake [2]. RNA extracted from liver, spleen, and intestines of the seven dead wild birds of three different species (3 Great Crested Grebes (Podiceps cristatus), 2 Black-headed Gull (Larus ridibundus) and 2 Spoonbill (Platalea leucorodia) were tested positive for the influenza A virus and for the H5N1 subtype in real-time RT–PCR.
One virus isolate from each species was cultivated in embryonated specific antibody-negative fowl eggs. The viruses were tested with a hemagglutination inhibition assay with reference antiserum and antiserum against viruses, previously isolated in Russia. The results showed close antigenic relationship between the investigated strains and those isolated at the Uvs Nuur Lake in 2009 as well as in the Russian Far East in 2008 (both clade 2.3.2). The strains A/black-headed gull/Tyva/8/2010, A/ spoonbill/Tyva/1/2010, and A/great crested grebe/Tyva/22/2010 were shown to be highly pathogenic in chicken (IVPI = 3) and mice (intranasal infection, MLD50=2.8 log10 EID50/ml) [8, 12].
We were able to observe neuropathological behavioral signs such as tremor and circular movements in mice infected with A/great crested grebe/Tyva/22/2010, a phenomenon not previously described for HPAI H5N1 viruses isolated in 2009. Affected mice were sacrificed and tissue samples were taken for morphological investigation as described by [11]. Numerous hemorrhages were detected in the internal organs. The heart muscle had multiple sites of necrosis. Muscle fibers were disorganized, and had multifocal leukocyte infiltration. We also found several sites of hyperemia in the brain (Figure 1). The first lesions in mice brain were detected on 1st d.p.i., and included swelling of blood vessels, petechial hemorrhages. On 3rd d. p.i there were observed multiple foci of necrosis of neurons and gliocytes, and activated microglia. On day 5 and on day 7 p.i. were found a significant increase of neuronal necrosis. By 10th d.p.i large areas of leukocyte infiltration in the cortex and local hemorrhage were also observed. In addition the neuronal nuclei had dark structures common to the cell nuclei disrupted by the virus [11]. As soon as we found some certain differences between the brain tissues of the infected and control groups we suggested that these disruptions can be related to the observed behavior signs.
The full-length genome sequence of A/great crested grebe/Tyva/22/2010 was determined and submitted to GenBank under accession numbers HQ131674 - HQ131681.
Phylogenetic analysis of the hemagglutinin (HA) gene showed a close relation to strains isolated during the outbreaks at the same location in 2009, at the Qinghai Lake in 2009 and in Mongolia in 2010 as all of them fall into clade 2.3.2 (Figure 2). We can presume that these strains originally appeared before or around 2009 at the Qinghai Lake and later in May 2010 caused an outbreak in Central Mongolia [6]. In June 2010 dead birds were found at the Uvs Nuur Lake and the viruses with 98-99% identity to Mongolian strains were isolated.
There is a limited data about migratory link between China, Mongolia, and Russia. It was shown that Bar-headed Geese (Anser indicus) and Ruddy Shelduck (Tadorna ferruginea) with attached GPS satellite transmitters migrated to the breeding grounds near H5N1 outbreak areas in Mongolia [13]. There is no certain data about the migration of bird species involved in the outbreak studied. But we can hypothesize that there is a presence of the migratory link through Central Asian Flyway. It is partially proved by our H5N1 virus phylogeny studies [7,14].
As Uvs Nuur Lake serves as one of the key stopovers on the flyway between South East Asia and Europe, early AIV detection in this area can play an important role in terms of outbreak prediction, early warning and isolating new AIV strains when they start spreading from Asia to Russia and Europe.
The strains related to ones described in this study, originally were isolated in the Qinghai Lake area from the following bird spices: great cormorants (Phalacrocorax carbo), brown-headed gulls (Chroicocephalus brunnicephalus), great black-headed gulls (Ichthyaetus ichthyaetus), greatcrested grebes (Podiceps cristatus), and bar-headed geese (Anser indicus) [15]. We can suggest three mechanisms of its appearance at Uvs Nuur Lake.
Figure 1: The cerebral cortex of infected mice: A- Control; B – 5 d.p.i.; C – 7 d.p.i. HE (hematoxylin and eosin)-stained sections. Black arrows – swelling; red arrows – necrosis; blue arrows – hemolysis.
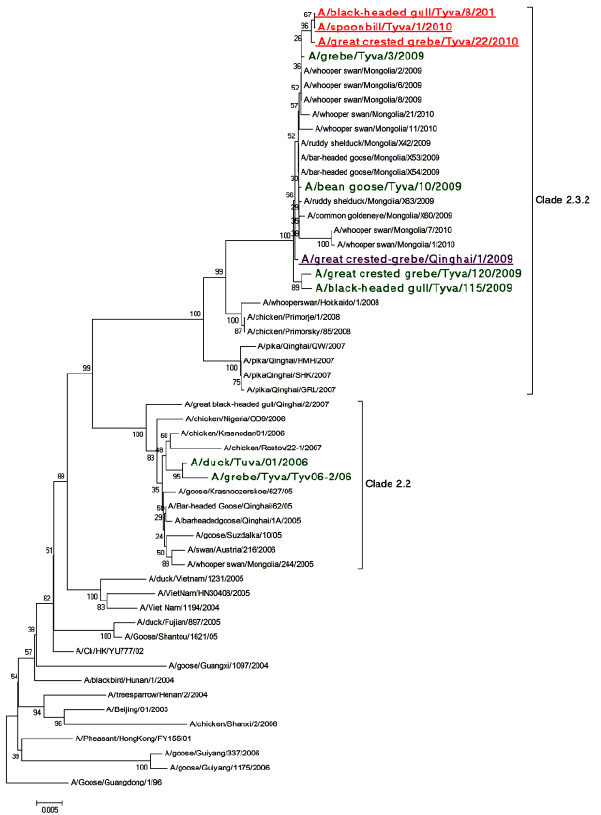
Figure 2: Bootstrapped (1000 replications) Minimum Evolution phylogenetic tree of HA genes. Viruses isolated at the Uvs Nuur Lake in this study are colored red and underlined; virus isolated in Qinghai Lake in 2009 is violet and underlined; viruses from wild birds previously isolated on Russian side of the Uvs Nuur Lake in 2006 and 2009 are green.
First is the direct transmission by the wild bird species, common for both areas. We think that grebes can play an important role in this mechanism. This species was involved in all detected outbreaks in both Uvs Nuur and Qinghai lakes since 2006 till now [3,7,15,16]. An important feature of Mongolia is that there is a little domestic poultry production in the country, therefore, AIV detection in wild birds would not likely be from the domestic poultry [17].
Second possible mechanism involves the virus transmission from one species to another along the flyway somewhere in between Qinghai and Uvs Nuur Lakes. The outbreaks in central Mongolia described by [6], can somehow confirm this hypothesis. HPAI viruses could be vectored by barheaded geese to the central Mongolia, where they had caused an outbreak and were also transmitted to grebes and gulls, which then vectored these viruses to Uvs Nuur Lake.
Another possible way is the establishment of local clade 2.3.2 HPAIV circulation at the Uvs Nuur Lake after its introduction in 2009 [7]. This circulation can involve some local animal species (birds, mammals) or the virus can be preserved in the environment [18, 19]. This hypothesis still needs further investigation.
Our experience with 2005-2007 outbreaks and isolated HPAI H5N1 (clade 2.2) viruses originally appearing around Qinghai Lake and then spreading first to Russia and later to Europe and Africa [1] leads us to the assumption of a similar scenario for clade 2.3.2 HPAI H5N1 virus spread. Recent detection of clade 2.3.2 H5N1 viruses during the recent outbreaks in Romania and Bulgaria [20], confirms this hypothesis.
Further investigation of the global distribution of clade 2.3.2 and 2.2 HPAI H5N1 viruses will give us better understanding of the Evolutionary Ecology of avian influenza viruses in natural host populations.
Download Provisional PDF Here
Article Type: Short Communication
Citation: Sharshov K, Susloparov I, Wang W, Li Z, Li L, et al. (2015) Neuropathological Effect of Clade 2.3.2 H5N1 Influenza Virus Isolated from Wild Birds. J Emerg Dis Virol 1(1): doi http://dx.doi.org/10.16966/2473-1846.102
Copyright: © 2015 Sharshov K, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access