Abstract
Studying the mechanism of bone defect healing is of great significance for targeted measures to promote bone defect healing. Macrophage, as one of the essential factors in the process of bone defect healing, can promote the bone repair by inducing bone formation and affect bone absorption. However, the upstream factors regulating macrophage to promote bone repair is unclear. Macrophage migration inhibitory factor (MIF) can regulate macrophage migration and phagocytosis, and is closely related to bone repair. This paper is to study whether MIF can regulate bone defect repair by macrophages. In this study, the expression and distribution of MIF during bone repair were detected by using rat drilling model. We found that MIF abundantly distributed in the lysosomes of macrophages, and was MIF+F4/80+ macrophages were distributed in regions where bone repair was active, such as the bone trabecular; Further, it was discovered that double positive cells in primary callus stage was much more than that in mature callus stage. These results indicated that MIF was involved in the process of bone defect healing promoted by macrophages, and the regulation of macrophage by MIF might be through lysosomes.
Introduction
Bone defect, caused by trauma, resection of bone tumor, osteomyelitis and congenital bone defect, causes great pain to the patients physically and psychologically. It is of great significance to study the specific mechanism of bone defect healing so as to take targeted measures to promote fracture healing. Macrophage is an indispensable factor in the process of bone repair. Raggatt, et al. [1] found that the mice bone could not be repaired with the macrophages being cleared. Macrophages promote bone repair by inducing bone formation and regulating bone resorption [2]. However, the factors regulate macrophages in bone repair is still unknown. MIF is an important pro-inflammatory factor, which can inhibit the migration of macrophages [3], promote the infiltration [4], aggregation, proliferation [5] and activation of macrophages in inflammation [6], enhance their adhesion and phagocytosis [7], and promote the production of a variety of inflammatory cytokines [8]. Considering the robust effect of MIF on macrophages, this study aim to observe whether MIF can promote bone repair by regulating macrophages.
Materials and Methods
Materials 12 SD rats of either gender
Provided by the Experimental Animal Center of the Fourth Military Medical University; holt’s acacia sucrose solution: obtained by 30g of sucrose and 1g of acacia dissolved in 100ml of ddH2O; OCT frozen section embedding agent (Tissue-Tek 4583); paraformaldehyde (IS014, Xi’an Hutter Biotech Co., Ltd); anti-fluorescence mounting medium containing DAPI (HNFD-02, Guangzhou Hao Chi Biotechnology Co., Ltd.); Anti-MIF antibody (ab7207, abcam, dilution factor 500X); Alexa Fluor 488 AffiniPure Donkey Anti-Rabbit IgG (H + L) - MIF corresponding secondary antibody (711-545-152, Jackson Immuno Research Inc, dilution factor 1000X); F4 / 80 (A-19) antibody (sc26642 , Santa Cruz, 300X); Alexa Fluor 594 AffiniPure Donkey AntiGoat IgG (H + L) - F4 / 80 corresponding secondary antibody (705- 585-003, Jackson Immuno Research Inc, dilution factor 1000X).
Rat tibia drilling model
12 rats were randomly divided into group A and group B, 6 rats in each group. Each group was randomly divided into experimental group and sham operation group, with 3 rats in each group. Group A was made for materials 7 days after operation and group B was made for materials 14 days after operation. Rats in the experimental group were injected intraperitoneally with 1% pentobarbital sodium (45mg per kg of body weight). The tibia was disrupted and shaved and disinfected at supine position, and was covered by the aseptic hole-towel. The disrupted skin of the tibia was cut by as calpel and the muscle attached on the media ledge of the tibia was cut by an eye scissor to reveal the tibia disruption. A drill of 2mm diameter was used to drill a hole in the tibia crest plane, through the unilateral cortical bone. The drill was washed by running’s a line water in the drilling process. The hole was washed by saline water once more. The skin was sutured with stylolite and disinfected by iodophor. The surgery ended there. The operation procedure of the sham operation group was the same as that of the experimental group, but without drilling.
Immunofluorescent staining
The acquisition of the frozen section of nerve tissue: The giant’s aphenous nerves and sciatic nerves were put into 4% paraformaldehyde immediately after obtain and fixed for more than 24hours. After that, they were dehydrated in Holt’s acacia sucrose solution for 72 hours. Then the nerves were embedded with the OCT frozen embedding medium in the freezing microtome (LEIKACM1850) at -20°C and sliced into 8um thick and preserved long-termined refrigerator at -20°C.
The acquisition of the frozen section of rat tibia drilling model: The rat used for the production of drilling model through operations was evacuated clear the blood by perfusion of 40°C saline water through blood vessel, followed by reperfusion of 4% paraformaldehyde. After that, the rat was fixed for dissection. The drilling area of the tibia was obtained and fixed in 4% paraformaldehyde for one more week. Then it was put into decalcification solution, which was changed every 3 days. The extra ends of the specimen were cut by a blade for the test of the decalcification state. After complete decalcification, the specimens were dehydrated in holt ‘s acacia sucrose solution for 72 hours and embedded with the OCT frozen embedding medium in the freezing microtome (LEIKA CM1850) at -20°C and sliced into 8um thick and preserved long-term in refrigerator at -20°C).
Immunofluorescent staining: The frozen sections were incubated at 37°C for 1h for rearming; PBS wash for 3mins × 3times; 0.25% Triton X-100 drilling for 30mins; PBS rinse for 3mins × 3times; 1% BSA blocking incubation at 37°C for 30mins; primary antibody incubation at 4°C overnight; PBS rinse 3 mins × 5 times; incubation with corresponding.
Second antibody added in at 37°C for 1h; PBS rinse 3mins × 5times; mounting after adding anti-fluorescence mounting medium containing DAPI. The result was observed by the included NISElement F software of Nikon Eclipse 80 microscope and the images were kept.
Results
Establishment of rat tibia drilling model and its postoperative healing status
We used the rat tibia drilling model to study the process of bone repair. HE staining showed that a large number of new bone trabecular were formed 7 days after operation, but all of them were relatively short. Bone trabecular began to connect along the stress direction 14 days after operation, the number of bone trabecular in the bone marrow cavity began to decrease, and the bone marrow cavity began to penetrate gradually. As shown in figure 1, bone repair is in the early stage of remodeling after 7d, and 14d is in the middle stage of remodeling.
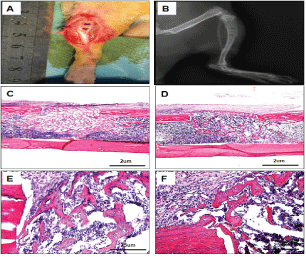
Figure 1: Establishment of a rat tibia drilling model and its postoperative healing state.
A. Operation diagram;
B. X-ray showed rat tibia drilling model;
C-F. HE staining showed fracture repair status;
C, E 7th days after operation;
D, F. 14th days after operation. HE staining showed that the fracture repair was in the early stage of remodeling and reconstruction at 7th days after operation, and was in the middle stage of remodeling and reconstruction at 14th days after operation.
MIF is phagocytosis by macrophages and localized in areas where bone repair activities is active
Immunofluorescence staining showed that F4/80 could label macrophages. MIF+F4/80+ double-positive cells were distributed in active bone repair areas, such as around new bone trabecular. In positive cells, lysosomes structure can be clearly displayed, and MIF was also localized in lysosome vesicles (Figure 2).
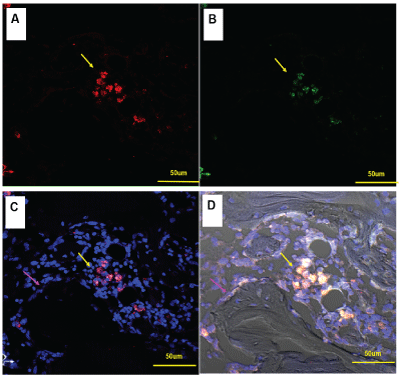
Figure 2: Distribution of MIF and macrophages in tibia drilling bone repair.
A-F4/80, red
B-MIF, green
B, C- DAPI –nucleus, blue
In positive cells (A), Lysosome structure can be clearly displayed, and MIF was also localized in lysosome vesicles (yellow arrow). The MIF + F4 / 80 + double positive cells were localized around the remodeling trabecular (purple arrow).
MIF+F4/80+double positive cells function more in the early stage of bone repair
Immunofluorescence staining showed that the number of MIF+F4/80+ double positive cells in bone defect is more on the 7th day than on the 14th day (Figure 3) after operation.
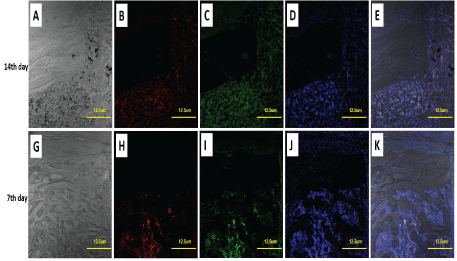
Figure 3: Distribution of MIF and macrophages in tibia drilling bone repair.
A-F4/80, red
B-MIF, green
B, C- DAPI –nucleus, blue In positive cells (A), Lysosome structure can be clearly displayed, and MIF was also localized in lysosome vesicles (yellow arrow). The MIF + F4 / 80 + double positive cells were localized around the remodeling trabecular (purple arrow).
Discussion
Macrophages are involved in every stage of bone repair, which requires removal of necrotic tissue [9] and elongation of new bone structures to restore normal bone structure and function [10]. Inherent immune cells, especially macrophages, can be detected immediately in the early stages of bone injury [11]. These macrophages induce a cascade of reactions that first remove the blood clots forming after injury and lead the m to be replaced by granulation tissues [12]; followed by the initiation of anabolism of granulation tissue, inducing its gradual evolvement into periosteal callus tissue or bone bridge available for the structure; these temporary bones structures were then gradually remodeled with normal bone structure and strength [13]. Macrophages are so important in bone repair, but the key factors that regulate macrophages in this process are not clear yet.
MIF plays an important role in bone repair. Mice with MIF-knock out suffer a serious delay in bone defects repair, and the mineral deposition rate in there pair process is significantly low compared with normal mice [14]. Onodera [15] et al., detected the expression and distribution of the MIF in mice femoral fracture model. Semiquantitative PCR showed that MIF was highly expressed throughout the healing process, reaching a peak on the fourth day after fracture. The immune histo chemical results showed that the increased expression of MIF was located in the active area of bone repair. MIF appeared in the granulation tissue 4days after fracture; appeared in thickened periosteal membrane, intramembranous osteoblast under it and the cartilage of the cartilage healing tissues 7-10 days later; and appeared in the cartilage of the cartilage healing tissues 14-28 days later. After that stage, the amount of MIF decreased gradually. This study showed that MIF+F4/80+ macrophages located in the bone remodeling active area, such as new bone trabecular formation area. Further, we found the double positive cells on the 7th day after operation was much more than that of 14th days after operation. These results demonstrate that MIF+ F4/80+ macrophages are involved in bone repair. And most of them function at the early stage of bone repair. In addition, MIF is more localized in macrophage lysosomes, suggesting that the regulation of macrophage by MIF may be through lysosomes. But the specific mechanism still needs further study.
Conclusions
MIF participates in the process of bone repair by macrophages. And its regulation of macrophage may be through lysosomes.
Article Information
Aritcle Type: Research Article
Citation: Li J, Wu H, Zhang SS, Jiang H, Wang J, et al. (2018) MIF Promotes Bone Defect Repair by Regulating Macrophages. Cell Stem Cells Regen Med 3(2): dx.doi.org/10.16966/2472-6990.119
Copyright: © 2018 Li J, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
Received date: 17 Aug, 2018
Accepted date: 05 Sep, 2018
Published date: 11 Sep, 2018
