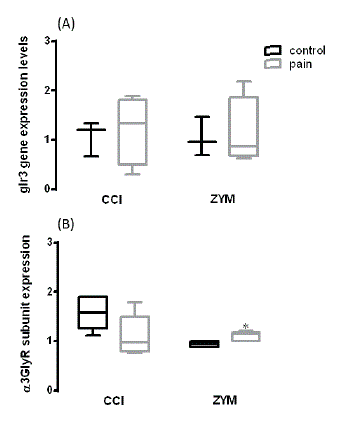
Figure 1: Differences in α3GlyR gene and protein expression in neuropathic and inflammatory pain. In (A) glr3 gene expression and in (B) is shown the protein expression.
Trinidad A Mariqueo*
Laboratory of Neuropharmacology, Center for Medical Research (CIM), School of Medicine, Universidad de Talca, Talca, Chile*Corresponding author: Trinidad A Mariqueo, Laboratory of Neuropharmacology, Center for Medical Research (CIM), School of Medicine, Universidad de Talca, Talca, Chile, Tel: 56-948875520; E-mail: tmariqueo@utalca.cl
Introduction: Increasing evidence suggests that changes in the balance of excitatory/inhibitory neurotransmission are involved in the development of the majority of chronic pain forms. In this context, impairment in glycine mediated inhibitory neurotransmission is thought to play a critical role in the disinhibition that accounts for the development and maintenance of central pain hypersensitivity.
Aims: The goal of this study was to evaluate the glycine receptor α3 subunit (α3GlyR) expression in neuropathic (Chronic Constriction Injury, CCI) and inflammatory (Zymosan A injected) animal models of chronic pain.
Results and Conclusion: RT-qPCR analysis of spinal cord samples showed that glra3 gene expression does not change after 3 days of CCI and 4 hours of Zymosan. A injection. However, we found that protein levels evaluated by Western blot increased after inflammatory pain. These data suggest that central sensitization is differentially regulated depending on the type of pain.
Central sensitization; Neuropathic; Inflammatory; Hypersensitivity; Glycine receptors
Chronic pain constitutes a major health problem, affecting millions of people worldwide. In the United States alone there are at least 116 million of adults affected by some form of chronic pain, representing an important medical and economic burden for the families of the individual affected [1]. Increasing evidence suggests that changes in the excitability of peripheral and central nociceptive pathways account for the development of the majority of forms of chronic pain [2]. In this context, impairment in GABA and Glycine mediated inhibitory neurotransmission are thought to play a critical role in the disinhibition that accounts for the development and maintenance of pain hypersensitivity [3]. Glycine is the main inhibitory neurotransmitter in the spinal cord and brain stem, playing a significant role in motor respiratory control and pain sensitization [4]. Glycine receptor (GlyR) subunits are encoded by glra and glrb genes [5]. In the mature spinal cord, glycine activates GlyRs, which are pentamers composed mainly of α1, α2, α3 and β subunits [6]. α3GlyRs are expressed in the dorsal horn spinal cord where the pain signaling is integrated [7]. It was reported that α3GlyRs under lies inflammatory pain sensitization by mechanisms that involve the Prostaglandin E2 signaling and the reduction of α3GlyRs activity [8,9].
Chronic pain comprises several different types of pain, including inflammatory pain following tissue injury (e.g. arthritis), cancer pain and neuropathic pain following nerve injury, spinal cord injury and brain injury (e.g. stroke and trauma) [10-12]. The pathogenesis of neuropathic and inflammatory pain involves the excitatory/inhibitory neurotransmission balance in a process related with the neuroimmune interface alteration [13-15]. Importantly, reduced inhibitory activity in the dorsal spinal cord has been reported in both persistent neuropathic [16-18] and inflammatory pain [19-21]. However, the information related with GlyR subunit gene expression in different types of pain it has not been precisely characterized.
Therefore, we evaluated the expression of α3GlyR subunit in neuropathic (Chronic Constriction Injury, CCI) and inflammatory (Zymosan A injected) animal models of pain sensitization.
The chronic constriction sciatic nerve Injury (CCI) and inflammatory hyperalgesia induced by subcutaneous Zymosan. A injection on the left hindpaw in Sprague Dawley rats are well known and extensively validated models of neuropathic and inflammatory pain [20,8]. Rats were maintained at the animal facility of the Universidad de Chile for about 4 weeks. Accordingly we used preferentially adult male animals because it was reported there exist sex differences in the modulation and expression of different types of channels involved in nociceptive processing [22-24]. All procedures conformed to the regulations specified by the Institutional Animal Use Committee of the University of Chile (n°17027-MED-UCH) and conducted according with the ethical protocols established by the National Institutes of Health, Bethesda, USA.
qPCR from spinal tissues of CCI rats were performed at 3 days postsurgery and were compared with data obtained from Sham (rat surgery without ligation). Quantitative PCR of spinal cord tissues derived from Zymosan-A-injected (0.06 mg in 20 μL PBS) and control rats (saline) were extracted 4 hours after hind paw Zymosan A injection. Total RNA was isolated from the spinal cord (L3-L5 lumbar segment) by the Guanidinium thiocyanate-phenol chloroform extraction method (Trizol, Life Technologies, USA). Following total RNA preparation from these tissues and cDNA synthesis, qPCR of α3GlyR subunit was performed using 0,5 ng/uL of cDNA and SYBR green Master mix (Sigma) in a final reaction volume of 20 uL in Stratagene Mx3000P equipment (Agilent Technologies, USA). PCR amplification cycles were: 1 at 95°C/5min, 40 at 95°C/30s, 56°C/30s and 72°C/45s. All reactions were run in triplicate. The relative expression of each gene was calculated using the Pfaffle Method [25] and normalized against GAPDH expression (Table 1).
| Primers | 5ˈ- 3ˈ Sequence | Amplicon (Base pairs size) |
| GAPDH-R | GGCCTCTCTCTTGCTCTCAGTA |
141 |
| GAPDH-F | TTGTGAAGCTCATTTCCTGGTA | |
| Alpha3-R | GCCTTCTGATTGTCATTCTGTC |
109 |
| Alpha3-F | CTCTGCGTGGTCATCGTAAG |
Table 1: Primer sequences used in qPCR experiments.
Total proteins were quantified using the Micro BCA protein assay (Pierce Biotechnology, Rockford, IL). SDSPAGE was performed using 60 μg of protein on NuPAGE gels (Invitrogen Life Technologies, Carlsbad, CA). Proteins were transferred to a PVDF membrane (BioRad, Hercules, CA) and then blocked overnight by incubation with 5% dry milk in Tris-buffered saline containing 0.1% Tween 20 (TBS). After washing with TBS, the blots were incubated with anti α3GlyR subunit (1:300, rabbit; Millipore) primary antibodies. Monoclonal anti-GAPDH antibody was used as load control.
All data are presented as mean ± standard error of mean (SEM). Statistical analyses were performed using graph pad prism 6 software. One-way ANOVA analysis was performed. P-value<0.05 was considered as significant.
To understand the contribution of α3GlyRs to the central sensitization process, α3GlyR subunit expression levels were analyzed in spinal cord tissues from neuropathic and inflammatory animal models of pain. In figure A, it can be seen that the glr3 gene expression levels did not change after 3 days of CCI. Similar results were obtained after 4 hours of Zymosan A injection. We evaluated the glr3 gene expression at this time because it was reported that after 3 days of CCI and 4 hours of Zymosan A there are significant changes in nociceptive threshold [20]. Both have been used before as animal models to study chronic and acute neuropathic-inflammatory pain [26,27].
Figure 1: Differences in α3GlyR gene and protein expression in neuropathic and inflammatory pain. In (A) glr3 gene expression and in (B) is shown the protein expression.
However, data show that α3GlyRs protein expression increased after inflammatory pain and no changes were observed after 3 days of CCI (Figure B). These results suggest that the central pain sensitization process is differentially regulated by glr3 gene expression. Previous studies have described that inhibitory glycinergic neurotransmission is altered in chronic pain conditions. In order to understand the contribution of α3GlyRs to central sensitization, Harvey RJ, et al. (2004) generated a Glra3-/- Knockout mice. Behavioral approaches demonstrated that pain sensitization induced by intrathecal PGE2 and peripheral inflammation was reduced in Glra3-/- Knockout mice [20]. In agreement with these results, Reinold H, et al. (2005) showed that mice deficient in EP2 receptors completely lack spinal PGE2 mediated hyperalgesia [21]. Electrophysiological experiments showed that α3GlyR mediated currents were inhibited by PGE2. These effects were abolished by intracellular loop mutation at serine 346 and PKA inhibition, demonstrating that phosphorylation of α3GlyRs consistently determines the inflammatory pain sensitization [26]. In agreement with these reports, our data show that the α3GlyRs protein increased only in the inflammatory context, suggesting the existence of fine tune mechanisms that regulate the protein expression in different types of pain conditions. In fact, Harvey VL, et al. in 2009, showed no differences in pain behavior of Glra3-/- Knockout mice in comparison with the control mice after neuropathic pain surgery [27]. Furthermore, other subunits of GlyRs have been related with the establishment of neuropathic pain. Imlach WL, et al. showed that α2GlyR subunit expression was increased in the dorsal horn of spinal cord after neuropathic pain [29]. We have evidence that regulatory beta auxiliary subunit of glycine receptors play important role in neuropathic pain (under publication). On the other hand, previous studies have demonstrated that the activation of GlyRs triggers the chloride ion influx, which results in the membrane hyper polarization needed to control the excitability of neural circuits. Chloride homeostasis is regulated by co-transporter KCC2 [29]. One of the critical steps in the central pain sensitization that accounts for disinhibition is related with the KCC2 down regulation [30]. It has been reported that KCC2 expression is altered in the central pain sensitization process, leading to the collapse of chloride ionic gradient and shifting the glycine receptor reversal potential to more depolarized values [31]. In terms of the molecular mechanism involved in this process, Coull JA, et al. (2005) showed that ATP-stimulated microglia produce a depolarizing shift in anion reversal potential in spinal neurons [17]. The authors conclude that molecular mechanisms involved in these changes are associated with the activation of TrkB receptors by BDNF because blocking these signaling with anti-TrkB antibody or siRNA anti BDNF abolished the changes in the anion reversal potential. On the other hand, Zhou HY, et al. (2012) demonstrated that a shift in glycine current reversal potential is mediated by KCC2 calpain cleavage [32]. A recent study showed that morphine tolerance is induced by the down regulation of KCC2, impairing the chloride homeostasis in rat spinal neurons [33]. How the expression of different GlyRs can be influenced by KCC2 changes is a question not addressed yet. Future studies are necessary to propose new therapeutics alternatives for the management of chronic pain.
Our results showed that α3GlyRs protein increased in inflammatory pain conditions, suggesting that central pain sensitization fine-tunes gene expression regulation in different types of pain contexts.
I thank professional assistance of Nicole Cespedes for collecting part of qPCR data, Gabriela Améstica and Nicolas Sepulveda for collecting Western blot data.
This work has been supported byPIEI QUIMBIO University of Talca (Mariqueo TA) and FONDECYT grant No. 3170690 (Mariqueo TA).
Author declares no conflicts of interest
Download Provisional PDF Here
Article Type: RESEARCH ARTICLE
Citation: Mariqueo TA (2020) The Expression of Glycine Receptor α3 Subunit is Differentially Regulated in Different Types of Pain. J Neurol Neurobiol 6(2): dx.doi.org/10.16966/2379-7150.161
Copyright: © 2020 Mariqueo TA. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access