Abstract
Objective: The prevalence of congenital urinary tract defects and congenital hydronephrosis is higher among males than females. Aquaporin (AQP)-1-3 are involved in the mechanism of pathophysiologic changes in the obstructed kidney. The aim with this study is to investigate whether gender differences affect the expression of AQP1-3 in response to partial unilateral ureteral obstruction (PUUO) for one week in newborn rats.
Materials and Methods: Forty-nine newborn rats were randomly allocated into 2 groups: PUUO (n=26, 13 males, 13 females) and Sham (n=23, 16 males, 7 females). The left kidney was subjected to PUUO within two days of birth. Sham rats were studied in parallel. After one week the animals were sacrificed and the kidneys were harvested.
Results: AQP2 and –3 mRNA and protein expression were decreased in male obstructed kidneys compared to Sham, consisting with a weaker labeling in the obstructed kidneys. In PUUO rats, AQP2 expression and labeling were reduced in male compared with females. AQP1 expression was lower in male compared with female rats on the contralateral non-obstructed side and this was consistent with a weaker labeling.
Conclusion: The present study demonstrated reduced AQP2 expression in males compared with females following PUUO. This impact of gender on AQP2 regulation might be due to the difference in sex hormones and glomerular hemodynamics.
Keywords
AQP1-3; Gender; Newborn rats; PUUO; Kidney
Introduction
Congenital obstructive nephropathy is the leading cause for renal impairment and a serious renal disease during infancy and childhood [1]. Approximately, 1% of all newborn infants are prenatally diagnosed as having hydronephrosis [2].
Interestingly, the overall prevalence of congenital urinary tract defects is 62% higher at birth among boys than among girls [3]. Especially, congenital hydronephrosis is 111% more frequent in boys [3]. In addition, the prevalence of congenital renal agenesis is higher in boys compared with girls (59.4% vs 40.6%) [4]. These findings indicate that gender may play a role in congenital kidney pathology. However, the underlying pathophysiological mechanisms are still unclear.
The consequence of obstruction is characteristically impairment of urinary concentration capacity. This involves two renal tubule segments: the loop of Henle and the collecting duct. The loop of Henle is in a large proportion responsible for generating the hypertonic medullary interstitium by countercurrent multiplication by NaCl active absorption in water impermeable nephron segments. The collecting duct is vasopressindependent osmotic equilibration of urine with the hypertonic interstitium
by the high water permeability (constitutive or vasopressin-regulated) mainly depending on aquaporins (AQPs) [5].
Aquaporins are proteins that facilitate water transport across biological membranes. In the kidney, AQP1 is predominately responsible for the constitutive water permeability of the proximal tubules and the thin descending limbs of Henle [6]. AQP2 and -3 are located in the principal cells of the collecting ducts and are regulated by vasopressin [6].
Previous studies have demonstrated decreased expression of AQP1-3 in rats subjected to PUUO for up to 14 weeks [5]. However, the impact of gender has not been elucidated. Our hypothesis is that male PUUO rats will have reduced expression of AQP1-3 compared to female PUUO rats. Therefore, the aim of this study was to investigate if gender has any impact on AQP1-3 expression in newborn rats subjected to PUUO for one week.
Materials and Methods
Animals
All procedures conformed with the Danish National Guidelines for care and handling of animals and to the published guidelines from the National Institutes of Health. The experimental animal protocols were according to the licenses for use of experimental animals issued by The Ministry of Food, Agriculture and Fishes, Animal Experiments Inspectorate (Approval no. 2010/561-1830).
The experiments were performed in 49 newborn Munich-Wistar rats (Møllegaard, Aarhus, Denmark) with an average weight of 5–8 g. The rats were subjected to PUUO or sham operation within the first 48 h of life as described previously [7], and randomized into two groups: PUUO (n=26) and Sham (n=23).
Newborn rats are poikilothermic and incapable of maintaining their body temperatures adequately. Partial ureteral obstruction on left side was performed according to a modification of the technique by Ulm and Miller [8]. The newborn rat was in deep sleep after placing on crushed ice for about 8 min, and then removed to the operating table. The left ureter was exposed through a midline incision, two-thirds of it was embedded in the underlying psoas muscle groove. Sham rats were performed by a laparotomy and mobilization of the left ureter. Following surgery, the rats were placed in an incubator maintained at 28°C until totally conscious and were then returned to their mother. The mother rats had free access to a standard rodent diet and were kept in an animal facility maintained at a controlled room temperature of 21 ± 2°C and a 12:12-h dark/light cycle. The rats were allocated into four sets of protocols, according to Table 1.
Gender determination
Tail clips were digested by lysis buffer with 200 mM NaCl, 100 mM Tris-Cl, 5 mM EDTA, 0.2% SDS and 10 mg/ml proteinase K overnight at 55°C. DNA was precipitated by isopropanol, washed by 70% ethanol and then resuspended in 100 µl TE buffer. Sry gene is a male-specific gene on the Y chromosome which is responsible for the male sex determination. Male gender was determined by amplification of a 317-base pair Sry sequence by PCR. Adult male and female rats were considered as controls. Sry gene was determined by PCR amplification with primer pairs (Table 2). PCR conditions were: 30 cycles denaturation for 30 s at 95°C, annealing and polymerization at 60°C for 1 min. PCR products were analyzed by electrophoresis in a 1% agarose gel at 70 V for 45 min.
RNA extraction and QPCR
Total RNA from rat tissue was isolated using the Machery-Nagel’s Nucleospin® RNA II kit (AH Diagnostics, Denmark). cDNA synthesis was performed with Fermentas RevertAidTM First Strand cDNA Synthesis Kit (MBI Fermentas, Ontario) according to the manufacturer’s instructions. PCR was performed using 100 ng cDNA as a template for PCR amplification. We used Maxima® Hot Star PCR Master Mix according to the manufacturer’s instruction (Thermo Scientific, Denmark). A standard curve was constructed by plotting threshold cycle (Ct values) against serial dilutions of cDNA. Primer sequences were used according to Table 2.
Western blot
Gel samples were run on a 12% Criterion stain-free gel (Bio-Rad, Denmark). After electrophoresis, the gel was activated using a ChemiDoc MP imaging system and the Imagelab software (Bio-Rad, Denmark). Proteins were transferred to a nitrocellulose membrane and blocked with 5% non-fat dry milk in PBS-T (80 mM Na2 HPO4 , 20 mM NaH2 PO4 , 100 mM NaCl, 0.1% Tween 20, adjusted to pH 7.4 using 10 M NaOH). After washing with PBS-T, the blots were incubated with primary antibodies overnight at 4°C (Table 3). The antigen–antibody complexes were visualized after incubation with horseradish peroxidase (HRP)-conjugated secondary antibodies (P448; dilution 1:4000; DAKO, Denmark) [9].
Immunohistochemistry
Kidneys were fixed by perfusion via right ventricle with 3% paraformaldehyde in 0.1 M cacodylate buffer, pH 7.4. Fixed kidneys were dehydrated in graded ethanol and left over night in xylene. The tissue was embedded in paraffin and cut into 2 µm sections on a rotary microtome. The sections were deparaffinized and rehydrated. For immunoperoxidase labeling, endogenous peroxidase were blocked by 0.5% H2 O2 in absolute methanol. To reveal antigens, sections were boiled in TEG-buffer with 10 mM Tris supplemented with 0.5 mM EGTA. Aldehydes were quenched in 50 mM NH4 Cl for 30 min, followed by blocking in PBS supplemented with 1% BSA, 0.05% saponin, and 0.2% gelatin, and then incubated overnight at 4°C with primary antibody (Table 3). After washing the sections were incubated with horseradish peroxidase-conjugated secondary antibody (P448, goat anti-rabbit immunoglobulin, DAKO, Denmark) Antigenantibody reactions were visualized with 0.05% 3,3’-diaminobenzidine tetrachloride (Kem-en Tek, Denmark). The light microscope was carried out with Lecia DMRE (Leica Microsystems).
Hematoxylin and eosin (HE) staining
HE staining was performed using a standard procedure. Tissue was de-paraffinized in xylene, rehydrated in EtOH. Nuclei were stained with hematoxylin, followed by cytosolic staining with eosin, and lastly dehydrated in EtOH. Sections were evaluated under a light microscope of Lecia DMRE.
Statistics
Mixed-model with pairwise comparisons of margins as post estimate analysis was used for statistical analysis (Stata 12). P<0.05 was considered to indicate a significant difference. Results are given as means ± SEM unless stated differently
Results
PCR of Sry sequence
PCR analysis of the Sry sequence generated a 317-bp product in the adult male kidney, but not in the female one, as a reference. The same product was observed in 29 rats, which were determined as male (Figure 1A). Here 16 were sham operated and 13 were subjected to PUUO. Of the females, 7 were sham operated and 13 were subjected to PUUO.
Hydronephrosis degree related to AQP2 mRNA level
The hydronephrosis had different degrees after PUUO. Some kidneys expanded markedly with upstream ureter dilation proximal to the embedding part and were designated as “Large”. Kidneys with slight enlargement were designated as “Slight”; kidneys with dilation degree in between were designated as “Medium”. As shown in Figure 1B, both male and female animals with a slight degree of hydronephrosis had an increased AQP2 expression. HE staining was performed to investigate renal morphology and the histological findings of the obstructed, contralateral and Sham kidneys showed no difference between genders among the three groups (Figure 1C).
The impact of gender on AQP1 expression in response to PUUO
In male animals, AQP1 mRNA expression was decreased in obstructed kidneys compared to contralateral and Sham kidneys. In female animals, AQP1 mRNA expression showed no significant difference among the three groups (Figure 2A). Western blot analysis showed a band at 29 kDa corresponding to the non-glycosylated form of the anti-AQP1 antibody (Figure 2B). In male rats, AQP1 protein was decreased in both the obstructed and contralateral kidneys compared to kidneys from shamoperated rats. Female rats showed no difference in AQP1 protein levels among the three groups. In the contralateral kidneys, AQP1 protein was significantly lower in male rats compared with females (Figures 2B and 2C). Immunohistochemistry showed localization of AQP1 in the apical membranes of the proximal tubule and thin descending limbs of Henle (Figure 2D). AQP1 labeling was weaker in both obstructed and contralateral kidneys compared with Sham in the male rats (Figures 2Da, 2Dc and 2De). In contralateral kidneys, AQP1 labeling was weaker in male compared to female rats (Figures 2De and 2Df).
The impact of gender on AQP2 expression in response to PUUO
In both male and female rats, AQP2 mRNA expression was significantly decreased in both obstructed and contralateral kidneys compared to Sham, respectively. Furthermore, male sham rats had a reduced AQP2 mRNA expression compared to female sham operated rats (Figure 3A).
Figure 3B showed 25 kDa bands representing the non-glycosylated form of AQP2. In male rats, AQP2 protein level was decreased in the obstructed kidneys compared with contralateral and Sham kidneys. In female rats, no change in AQP2 protein level among the three groups was observed. In the obstructed kidneys, AQP2 protein level was decreased in the male rats compared with female rats (Figures 3B and 3C).
Immunohistochemistry showed detection of AQP2 in the principal cells of the collecting ducts (Figure 3D). Consistent with Western blot analysis, male rats showed weaker labeling of AQP2 in obstructed kidneys compared with contralateral and Sham kidneys, respectively (Figures 3Da, 3Dc and 3De. In addition AQP2 labeling was weaker in the obstructed kidneys from male compared with female rats (Figures 3Dc and 3Dd).
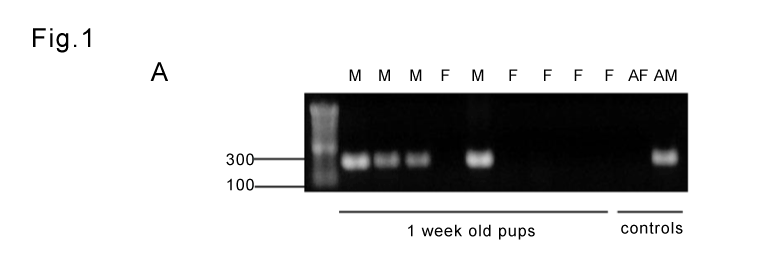
Figure 1A: Representative figure showing PCR gender typing of 1-weekold
newborn rats. The detection of the 317-base pair PCR product of the
Y-chromosome-specific gene indicates male (M) gender. The absence
of the Sry product indicates female (F) gender. An adult male (AM) and
an adult female (AF) were used as control.
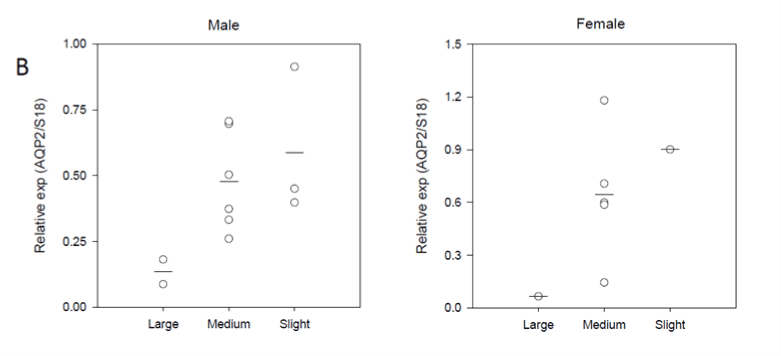
Figure 1B: Relationship between kidneys size from obstructed kidneys
and AQP2 mRNA level (normalized by housekeeping gene S18) showing
that AQP2 mRNA was inversely proportional to the kidney size.
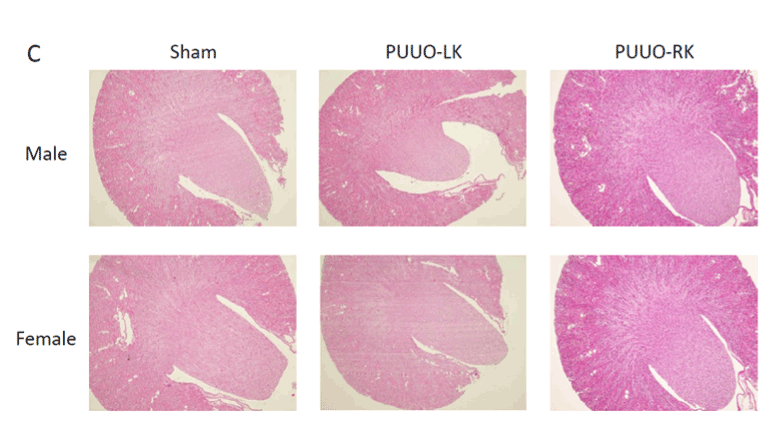
Figure 1C: HE staining of obstructed, contralateral and Sham kidneys
from male and female rats.
The impact of gender on AQP3 expression in response to PUUO
In male rats, AQP3 mRNA expression was decreased in obstructed kidneys compared to Sham. However, female rats demonstrated reduced AQP3 expression in both obstructed and contralateral kidneys compared with Sham (Figure 4A). The anti-AQP3 antibody identified bands at 29 kDa corresponding to the non-glycosylated forms (Figure 4B). In male animals, AQP3 protein level was decreased in obstructed kidneys compared to Sham but there was no change in female rats between the three groups. There was no difference in AQP3 mRNA and protein levels between genders in each group (Figures 4A-4C).
Immunohistochemistry revealed localization of AQP3 in the basolateral membranes in the principle cells in the collecting ducts. In male rats, AQP3 labeling was weaker in obstructed kidneys compared to Sham (Figures 4Da and 4Dc). However, no change in the staining intensity of AQP3 was observed among the groups in female rats (Figures 4Db, 4Dd and 4Df) and AQP3 labeling between genders showed no difference in each group (Figure 4D).
Discussion
To our knowledge, this is the first study investigating the impact of gender on the regulation of renal AQP1, -2, and -3 in both obstructed and
contralateral non-obstructed kidneys after one week PUUO in newborn
rats. Obstruction caused a decrease in AQP2 and -3 expressions compared
with Sham which only occurred in male rats. Moreover, AQP2 expression
was significantly decreased in male obstructed kidneys compared to
females whereas no effect on AQP1 and -3 expressions were observed.
We noticed different hydronephrosis degrees in both male and female rats. This phenomenon may be a result of the growth of obstruction within the animal because the kidney was still during development within 2 weeks after birth or the surgery itself i.e. the different proportion of embedded ureter length. However, even in grown out animals with very standardized obstruction a large variation in hydrophrosis degree is observed. Nephrogenesis is complete before birth in human beings but not in rats. Only 10% of the nephrons are mature when rats are born and nephrogenesis is not complete until after the first two weeks after birth [10]. PUUO that mimic upper urinary tract dilation in human second and third trimester fetuses, can be obtained by embedding the ureter in an incision in the psoas muscle using newborn rats within two days after birth.
Gender differences in renal diseases have been reported both in animal and human studies [11,12]. In chronic renal disease resulting from a number of different etiologies, such as in polycystic kidney disease, membranous nephropathy, immunoglobulin A nephropathy, and “chronic renal disease of unknown etiology” men progress at a faster rate to end-stage renal failure than women [13]. Several other types of renal disease progress at a faster rate in men compared with women, but the reasons for this sex difference are not well understood. It is tempting to speculate that gender-related differences in renal mass or nephron number, systemic or glomerular hemodynamics, and the direct cellular effects of sex hormones could play a role in sex differences. Although kidney weight and size are larger in male than in female in both animal and human [14,15], glomerular filtration rate (GFR) is not different between genders after correction for kidney weight or body surface area [16,17]. However, the glomerular hemodynamic adaption to stress may be different between sexes. Renal injury induced by neonatal PUUO is associated with reductions in GFR and reduced AQP2 expression [18]. Moreover, it has been demonstrated that testosterone could exacerbate renal dysfunction leading to reduced GFR in response to 3 days UUO [19]. Our data showed reduced AQP2 expression in male obstructed kidneys compared with females and one could consider that testosterone could play a potential role in the down regulation of AQP2 in male compared to female after renal obstruction. Previous studies have shown that female sex hormones, including estrogen is able to alleviate the progression of kidney disease [20]. In addition, estrogen may preserve the split renal function (GFR) in rats subjected to complete obstruction [21], indicating that estrogen might be involved in the progression of obstructive nephropathy. Expression of AQP2 by endometrial cells has been shown to correlate with estrogen levels in women and a recent study identified an estrogen response element in the AQP2 gene promoter [22], providing a molecular basis for sex hormone regulation of AQP2. Previous studies found that reduced AQP2 expression was restored to control levels after estrogen administrated in the urinary bladder from ovariectomized female rats [23], indicating that estrogen might play a role for the regulation of AQP2. Whether estrogen might be involved in the gender differences in the regulation of AQP2 in response to PUUO remains to be determined. However, we have previously demonstrated reduced AQP2 expression in PUUO rats which is prevented by the angiotensin-II type 1 receptor (AT1R) antagonist candesartan [18]. This suggests that blockade of AT1R may protect the neonatal obstructed kidney against development of obstructive nephropathy. A potential interaction between estrogen and AT1R has become an important matter of intense investigation because of the observed relationship between sex difference and the development of angiotensin II-associated renal diseases. Studies have linked the presence of estrogen with down regulation of AT1R activity and increased AT1R activity after ovariectomy could be prevented by estrogen replacement. Furthermore, renal AT1R binding is significantly greater in male than in female in normal condition [24]. Together with our findings, this could support the idea that estrogen could attenuate AQP2 reduction in response to PUUO in female rats via tonic inhibition of the AT1R. Vasopressin interacts with the vasopressin V2 receptors (V2R), which results in increased water permeability through the insertion of AQP2 into the apical membranes of principal cells in the collecting ducts [25]. Liu J et al. [26] found that V2R expression was increased in female compared to male rats. Previous studies from our laboratory have demonstrated that V2R expression is decreased in male post obstructive kidneys together with a marked down-regulation of AQP2 [27]. It could be speculated that an increased V2R expression might exist in female PUUO rats which could contribute to the increased AQP2 levels. We did not observe any gender differences in the expression of AQP1 and -3 in rats subjected to PUUO. Carol et al. [28] reported that the renal AQP1 expression was higher in males than in females in adult rats, but there was no significant difference between genders in prepubertal rats. This indicated that the male-dominant AQP1 expression developed after puberty, and might be the reason why there is no impact of gender on AQP1 in newborn rats. Consisting with our data it has previously been shown that there was no gender difference of renal AQP3 expression in adult mice [29]. The underlying pathophysiological mechanisms of the impact of gender on AQP3 expression in the developing kidney needs to be studied in further experiments. In conclusion, the present study demonstrated that AQP2 expression was reduced in males compared with females following PUUO. This impact of gender on AQP2 regulation might be due to the difference in sex hormones and glomerular hemodynamics.

Figure 2:The impact of gender on AQP1 expression in PUUO rats:
A: Quantification of AQP1 mRNA expression in obstructed, contralateral and Sham kidneys from male and female rats. Data are means ± SEM. Statistics (mixed models): a:b, c:d, e:f, a:e, b:d:f, non-significant and a:c, c:e, P<0.05.
B: Western blotting performed for protein level of AQP1 and GAPDH in obstructed, contralateral and Sham kidneys from male and female rats.
C: Densitometric analysis of protein band intensity for AQP1 relative to total protein in rat kidneys. Data are means ± SEM. Statistics (mixed models): a:b, c:d, b:d:f, non-significant and a:c, a:e, e:f, P<0.05.
D: Immunohistochemical staining of AQP1 in cortex in obstructed, contralateral and Sham kidneys from male and female rats. AQP1 is localized in the apical membranes of the proximal tubules.
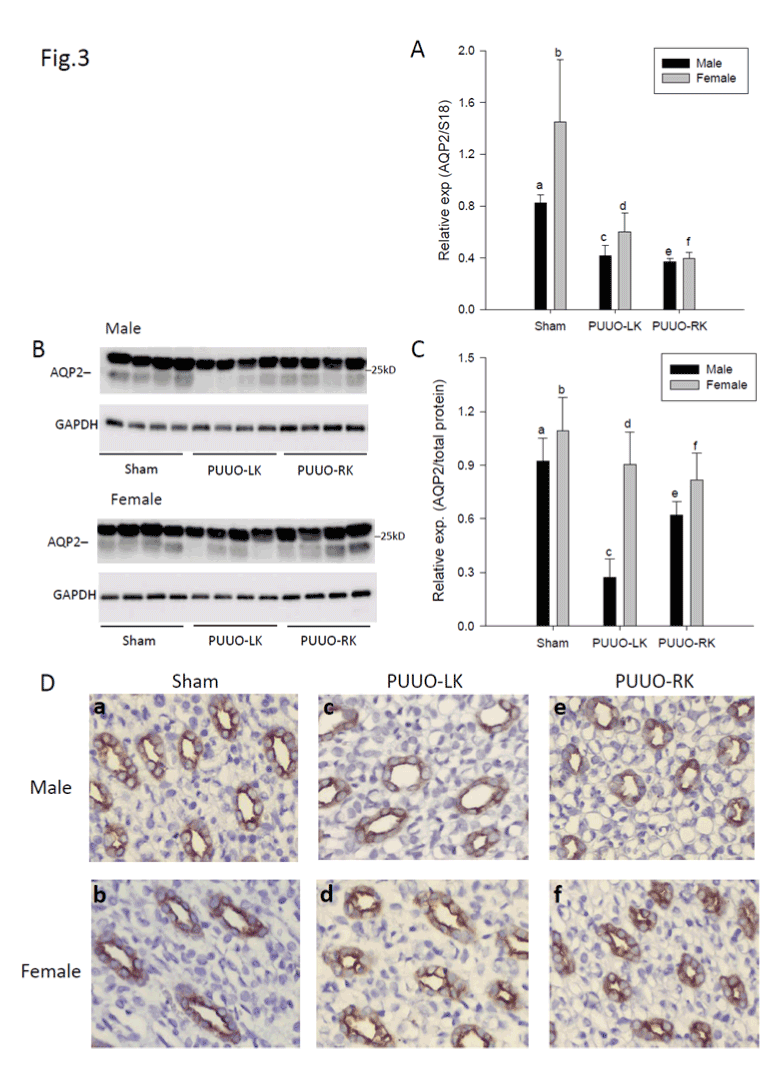
Figure 3:The impact of gender on AQP2 expression in PUUO rats:
A: Quantification of AQP2 mRNA expression in obstructed, contralateral and Sham kidneys from male and female rats. Data are means ± SEM. Statistics (mixed models): c:d, e:f, c:e, d:f, non-significant and a:b, a:c, a:e, b:d, b:f, P&0.05.
B: Western blotting performed for protein level of AQP2 and GAPDH in obstructed, contralateral and Sham kidneys from male and female rats.
C: Densitometric analysis of protein band intensity for AQP2 relative to total protein in rat kidneys. Data are means ± SEM. Statistics (mixed models): a:b, e:f, a:e, b:d:f, non-significant and a:c, c:e, c:d, P<0.05.
D: Immunohistochemical staining of AQP2 in medulla in obstructed, contralateral and Sham kidneys from male and female rats. AQP2 is localized in the principal cells of the collecting ducts.

Figure 4: The impact of gender on AQP3 expression in PUUO rats:
A: Quantification of AQP3 mRNA expression in obstructed, contralateral and Sham kidneys from male and female rats. Data are means ± SEM. Statistics (mixed models): a:b, c:d, e:f, c:e, non-significant and a:c, b:d, b:f, P<0.05.
B: Western blotting performed for protein level of AQP3 and GAPDH in obstructed, contralateral and Sham kidneys from male and female rats.
C: Densitometric analysis of protein band intensity for AQP3 relative to total protein in rat kidneys. Data are means ± SEM. Statistics (mixed models): a:b, c:d, e:f, a:e, c:e, b:d:f, non-significant and a:c, P<0.05.
D: Immunohistochemical staining of AQP3 in medulla in obstructed, contralateral and Sham kidneys from male and female rats. AQP3 is localized in the basolateral cells of the collecting ducts.
Acknowledgements
The expert technical assistance from Gitte Kall, Gitte Skou and Line V. Nielsen is highly appreciated. The authors thank Bo Martin Bibby for statistics help and Jens Christian Djurhuus and Jørgen Frøkiær for the support of the study