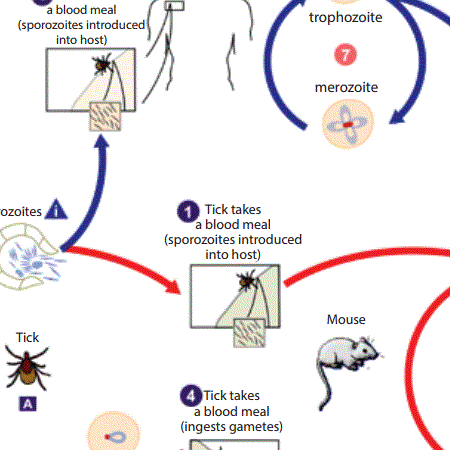
Figure 1: Babesia parasites life cycle.

Paulo Sérgio Balbino Miguel1 Andréia Patrícia Gomes1 Clayton Israel Nogueira1 Adriano Simões Barbosa Castro1 Paula Dias Bevilacqua2 Luiz Alberto Santana1 Lisa Oliveira3 Mauro Geller4 Rodrigo Siqueira-Batista1,5*
1Laboratório de Agentes Patogênicos, Departamento de Medicina e Enfermagem, Universidade Federal de Viçosa, Viçosa, Brasil*Corresponding author: Rodrigo Siqueira Batista, Departamento de Medicina e Enfermagem, Universidade Federal de Viçosa (UFV), Viçosa (MG), Brasil; Curso de Medicina, Faculdade Dinâmica do Vale do Piranga, Ponte Nova (MG), Brasil, Tel: (31) 3899-3978; E-mail: lmecs@ufv.br
Babesiosis is an infection caused by protozoa of the Babesia genus, whose vector are ticks of the Ixodes scapularis species. The disease is endemic in the United States, and, less frequently, in Europe; however, other regions occasionally may be affected. In non-endemic regions, like Brazil, the most of cases are associated with endemic areas travel. Thereby, the diagnosis should be proposed for patients with suggestive infection symptoms. Therefore, this review describes the illness caused by Babesia spp. and emphasizes the travelers’ health aspects. The discussion was based on the Cochrane, Lilacs, Pubmed, Scielo, and Scielo Brasil databases, including the citations available up to May 30, 2017. Traveler awareness about the possible transmittable illnesses at the destination, specifically babesiosis in the United States and Europe is fundamental.
Babesiosis; Protozoan infections; Traveler health
Babesiosis is an intraerythrocytic infection caused by protozoa of several species included in the genus Babesia. The pathogen is capable of infecting several animal groups naturally, including mammals, birds, reptiles, and amphibians [1,2] and humans are considered accidental hosts [3]. Transmission occurs by the bite of ticks belonging to the Ixodidae family, previously infected by the protozoa. In addition to this form of transmission, there are also cases caused by blood and congenital pathways [1,2].
In the United States and Europe, human babesiosis is generally associated with infection by Babesia microti and Babesia divergens, respectively [3]. However, Babesia venatorum emerged as a cause of the disease in China [4]. The first record of infection in the Homo sapiens sapiens species occurred in Yugoslavia in 1956 [5], and only three decades later Brazil reported the first case of the disease, more specifically in the state of Pernambuco [6], with further cases after this one uncommon in the country. However, the number of occurrences of human ixodidiosis has been increasing, accompanied by reports caused by agents transmitted by ticks [7,8].
The disease constitutes an occupational hazard for farmers, rangers, landscapers, hunters, and professionals who work in direct contact with the soil, road and railroad maintenance, and lumberjacking [9]. Travelers are of concern for health authorities in terms of babesiosis when considering the destination and duration of travel and the risk of exposure to infection [10]. It is important to point out that currently more than half a billion people are involved in international travel annually, which demonstrates the individual and collective risks arising from the movement of people and their interaction with the various environments [10,11]. Noting the zoonotic and economic importance of babesiosis-with a special focus on travelers’ health-this review presents the principal aspects related to etiology, life cycle, epidemiology, and clinical aspects of illness caused by Babesia spp.
Protozoa of the genus Babesia are included in the phylum Apicomplexa, of the suborder Piroplasmida and family Babesiidae [12]. They measure between 1 and 5 micrometers, and are oval, round or pyriform in shape. They present annular conformation and peripheral location, similar to the protozoa of the genus Plasmodium falciparum, which sometimes complicates diagnosis and may result in misidentification [13]. The absence of hemozoin deposits on the ring, absence of banana-shaped gametocytes, and presence of tetrads distinguish Babesia from Plasmodium [14].
More than 100 species infect several wild and domesticated animals and few have been confirmed as causative agents in humans, which vary depending on the region, with Babesia divergens, in Europe (especially in France and England), B. microti in the United States [15], more recently B. venatorum in China [4], also previously reported in Europe [16]. Cases of infection by Babesia duncani were registered in California and in the state of Washington [17]. Although, B. microti is present in Europe, there are few cases caused by this agent on the continent, most likely because the vector (Ixodes trianguliceps) is not a human parasite [18].
The Babesia species are classified into two forms, small and large. The former, represented by Babesia microti, Babesia duncani, and Babesia conradae, vary between 1 and 3 µm in diameter. Their trophozoites divide by two successive binary fissions, resulting in four merozoites, arranged in a tetrad [13,19]. The large forms include Babesia bovis, Babesia canis, and Babesia odocoilei, with diameters varying between 3 and 5 µm [13] and division only occurring in two pairs of merozoites (“paired form”) [13,19].
In vertebrates, including humans, the most common form of transmission of Babesia spp. is by the bite of ticks belonging to the Ixodidae family [2,13]. The endemism of the vector Ixodes scapularis, in Canada and most Eastern states of North America [20] coincide with that of babesiosis caused by B. microti [14].
The life cycle of I. scapularis is made up of three stages, larva, nymph, and adult [14]. The infection of newly hatched vector larva by Babesia spp. normally occurs at the end of summer. The parasite may cross the intestinal epithelium of the vector, with successive migration by the hemolymph and salivary glands [20]. However, the protozoa remain dormant during the transition from the larval to the nymph phase. At the start of the following summer, during the tick bite-together with the saliva of the nymph infected by B. microti-injection of sporozoites occurs, which are released on the host skin. All developmental forms – larva, nymph, and adult – feed on human blood; however the nymph is the principal vector due to its small size and greater activity in the summertime [14].
Transmission requires 36 to 72 hours, given that the sporozoites are only generated after activation, because of dormancy and exposure of the tick to the host’s warm blood, and are not immediately available in the salivary glands [14]. Afterwards, the sporozoite penetrates directly into the erythrocytes (B. bovis and B. divergens) or initially in the lymphocytes, where replication and release of “daughter cells” occurs, which in turn invade the red blood cells (B. microti). The host cell membrane undergoes intussusception, forming a vacuole that gradually disintegrates. Upon penetrating the erythrocyte, the organism transforms into the trophozoite form, able of moving freely in the cytoplasm [13].
The next step in the cycle is binary division, asexual reproduction known as merogeny. Depending on the type of trophozoite, there will be division into two or four merozoites with in the red blood cells [13,21]. The red blood cells lose integrity after the exit of the merozoite and the departure of this developmental form allows for the cycle to re-begin. In the blood, some trophozoites differentiate into male and female gametocytes, within the red blood cells [13].
During the tick’s blood meal, trophozoites, merozoites, and gametocytes are ingested. However, only the gametocytes survive the intestinal lumen of the tick, where sexual reproduction occurs. This is followed by penetration of the zygote into the cells of the intestinal endothelium and asexual reproduction (sporogony), which results in sporokinets. These reach various organs within the tick’s body by means of the hemolymph, including the ovaries and salivary glands, locations where sporogony occurs again, producing infective sporozoites that are inoculated together with saliva during the vector’s feeding [22]. Some species of Babesia spp. show transovarian transmission to vector for all phases of the life cycle [12].
Transmission of the protozoan to humans can also occur by congenital route or by blood transfusion [2,13]. The parasite can survive for up to 35 days at 4o C [23].
Transmission of Babesia sp. to humans occurs most frequently by the bite of the infected vector, but also occurs by blood transfusion. In this model (Figure 1), ticks infected by Babesia sp. introduce the sporozoites into the host during the bite, together with saliva (1). The sporozoites entre the red blood cells and reproduce asexually (2). In the blood, some parasites differentiate into male and female gametes (3). Upon ingestion by the tick, the gametes (4) unite forming new sporozoites (5). Humans enter the cycle accidentally, upon being bitten by ticks infected with Babesia sp., and consequent sporozoite injection (6), which penetrate the erythrocytes (B) and reproduce asexually (7). Blood transfusion is another form of transmission (8). Transovarian transmission occurs in the “large” forms of Babesia sp (A) (Source: Centers for Disease Control and Prevention (CDC)).
Figure 1: Babesia parasites life cycle.
Babesia divergens is the babesiosis cause, especially in Europe, and the transmission takes place via Ixodes ricinus, a tick with a three-year life cycle (larva, nymph, and adult). Most infections coincide with warmer weather (period of greater tick activity) and because of the higher number of people in areas of infestation [20].
Because of their peripheral position on the red blood cells, the parasites may be frequently confused with Plasmodium falciparum [13]. However, the species that cause babesiosis lack a tissue stage and rarely trigger large scale hemolysis, since their asexual reproduction is asynchronous [13,21].
Babesiosis is an emerging disease, widely distributed throughout the world, affecting several species of vertebrates, with the greatest impact on bovines and humans. Although humans are considered accidental hosts, the infections rates of this disease have increased considerable over the years [13,24,25]. In this sense, knowledge of the etiological agent, the vector, and the clinical characteristics are important for (1) clinical recognition of the disease (especially in terms of traveler’s health), (2) correct diagnostic approach, and (3) appropriate therapeutic management.
The disease is considered endemic in the US and in Europe; however, there are reports of occurrence in other regions. One of the more recent studies demonstrated 48 cases in China, most among women with an average age of 45 year, and the infection was caused by B. venatorum [4]. Obviously, these reports do not compare to those registered in the US, where from 2005-2010 more than 1400 cases of human babesiosis were reported [8], and in 2013, 1762 cases were reported in 27 states [9]. Because of its geographical distribution and the annual number of cases, babesiosis caused by the agent B. microti is currently considered to be an emerging disease, and the cases involving B. divergens are rare [13]. In the US, the zoonosis involves rodents of the Peromyscus leucopus species as reservoirs and intermediate hosts for the pathogen [13]. In Europe, the diseases more rare, but also more lethal, the most caused by B. divergens in splenectomized patients [9].
The description of babesiosis in humans on the European continent includes countries such as Scotland and Russia – especially associated with cattle rearing [2,13,26,27], in addition to Germany, Austria, Spain, Ireland, Italy, Switzerland, Sweden, and especially France and England [17]. Although sporadic, the disease also occurs in countries in Asia including South Korea, Japan, and Taiwan, and also in countries of the African continent [13]. The first case of babesiosis in Australia was reported in 2012, after microscopic examination of the blood sample from a 56 year old, who died in 2011. The samples demonstrated the presence of intra-erythrocyte parasites [3]. Other human infections caused by Babesia spp, also occurred in South Africa, in Egypt, and in Mexico, which may mean that this parasite has a much wider distribution than is currently known [17].
In Brazil, one of the cases of human babesiosis, whose diagnosis was not conclusive during the course of the illness with relapses, was identified in the city of Rio de Janeiro (RJ), and the patient subsequently died. This was a case of a splenectomized patient [8]. Brazil has also been cited as origin of a case of babesiosis, described in Poland in 1997 [28].
Babesiosis is often compared to malaria in terms of physiopathogenesis, due to the intra-erythrocyte location of the agents [13,29,30]. In fact, erythrocyte invasion is an important part of the Babesia spp. cycle, and it is very efficient at entering these cells in order to evade the human host response. Interaction between this pathogen and the host cell is very specific, since invasion occurs only in red blood cells, a fact attributed to the merozoites that are capable of locating, binding, and invading them [31].
This specificity is possible due to the multiple adhesive interactions that occur between the ligands present on the protozoan membranes and the target receptors the surface of the host cell [32]. In B. bovis, for instance, several genes code for merozoite surface antigens (MSAs), specifically MSA-1, MSA-2b, and MSA-2c [33]. This genetic diversity may be strategic to the survival of these protozoa [34], and the antigenic variations arise from these diversity profiles [33].
The host immune response is required for control of the infection caused by Babesia spp., important in this process are the cytokines, highlighting that the sequential expression of their genes can confer protective immunity [35]. Studies have also suggested action of the TCD8+ lymphocytes and natural killer cells in response to the disease [36]. In response to infection, there is leukocyte activation, production of tumor necrosis factor-alpha (TNF-α), and of interleukin 1 (IL-1) interleukin 2 (IL-2), and interleukin 6 (IL-6) [13,36], together with increasing levels of TNF which are, however, lower than those reached in malaria [37]. There is also lymphocyte activation and hypergamma globulinemia [38]. Specifically for B. microti, during the acute phase of infection, levels of IL-12, IFN-γ, and TNF-α are elevated and are crucial to control the initial explosion of parasite multiplication. In these patients, the most common complication is lung infection, with detection of IFN-γ within and around pulmonary blood vessels and TNF-α in the alveolar septum [35].
The function of the spleen in the disease pathogenesis is not completely established; however, the most severe form has been observed among patients with functional or anatomic asplenia. In part, the splenomegaly is associated with proliferation of phagocytic cells [13,39]. It is thought that the red blood cells infected by B. divergens alter spleen histopathology and cause alterations in the cell cycle and induce oxidative stress in this tissue [40].
Merozoites exit the red blood cells by rupture of the host cell, resulting in hemoglobin release in the blood stream. The free hemoglobin is immediately bound to haptoglobin and processed by phagocytic cells [13].
In general, the incubation period for babesiosis varies from one to three weeks, though it can extend to six weeks after the tick bite, or even up to nine weeks in case of infection by blood transfusion. On average, the time between symptom onset and diagnosis is 15 days. Among the cases verified in the United States and in Europe, the clinical manifestations differ markedly, considering that the infections are caused by different parasites [13].
The severity of the infection may vary with age and host immune status, simultaneously with other pathogens, and/or genetic factors. Acute infections in asplenic or immunocompromised patients, newborns, and the elderly, may give rise to complications, among which are: severe hemolysis, hemodynamic instability, acute respiratory failure, and multiple organ dysfunction syndrome, which may be fatal [41].
Symptoms are initially non-specific: malaise, fatigue, anorexia, fever (temperature above 38.0° C and relative bradycardia), headache, chills, joint and muscle pain, and behavioral changes with emotional lability or depression [13,41]. Fever, important on physical examination, may be constant or intermittent. Other symptoms cover the progression of the disease with acute respiratory distress syndrome (ARDS) [42], shock, petechia, and other hemorrhagic phenomena, and hepatosplenomegaly. Association with Lyme disease is not uncommon [13]. Also common are: discrete hepatomegaly and splenomegaly, and absence of lymphadenopathy. On computerized tomography, splenic infarcts and ruptures have been observed in the absence of palpable splenomegaly [13]. However, asymptomatic infection has been registered in roughly 20% of adult patients and 50% of pediatric patients, in a study spanning one decade in a highly endemic area [14].
In Europe – where most cases are caused by B. divergens – evolution is fulminant, with fever, prostration, anemia, and hemoglobinuria, after the three week incubation period. Hemoglobinuria is present, followed by jaundice, persistent fever (between 40.0 to 41.0° C), headache, chills, sudoresis, and myalgia [13,41]. In contrast, in the US, most infections registered are asymptomatic or mild, and in 30% of cases is related to asplenia, with a low mortality rate. The incidence of subclinical infections is high, however, the clinical alterations are more common in asplenic patients, patients with Lyme disease (Ixodes scapularis, also the vector that transmits Lyme borreliosis), the elderly, and the predisposed, such as HIV-positive patients and those with lymphoma [13,43-45]. Borrelia burgdorferi and Babesia microti infections are common and can cause more severe human disease [46]. The convalescence period usually lasts a few weeks, but can extend as long as eighteen months [38,47].
Diagnosis of babesiosis is usually established by blood smear, with Giemsa (or a derivative) staining [48], or by thick blood smear [13]. Detection of the pathogen, however, is obtained by visualization of the characteristic intra erythrocytic ring [47] (Figure 2), and the morphological similarity of these rings with those of Plasmodium spp. must be taken into account [20]. This fact highlights the importance of patient history, especially travel history to areas considered endemic [14]. However, confirmative tests based on molecular or serologic methods may confirm diagnosis [48], including in situations of low parasitemia [9]. Alternatively, in these situations, multiple smear tests should be performed over several days to confirm diagnosis [13].
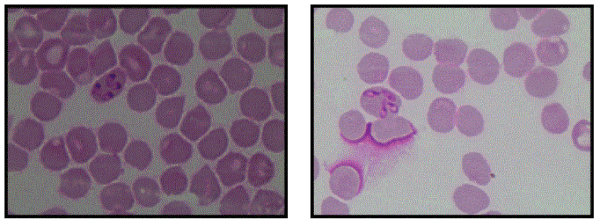
Figure 2: Babesia parasites in red blood cells on a stained blood smear (Aécio Carlos de Oliveira / UFV Veterinary Department).
The parasitemia of Babesia may also be obtained by inoculation of suspected patient’s blood into laboratory animals, easily detected when the parasite is B. microti [49]. Although the majority of routine laboratories do not have adequate structures for use of molecular and serological methods for rapid and reliable detection of Babesia, they should be considered for the diagnosis of the disease [50]. Among the most widely known molecular approaches is polymerase chain reaction (PCR), used in situations where parasite detection is difficult, in patients presenting symptoms suggestive of the disease [13]. Upon using primers for amplification of specific gene regions such as 18S rRNA and the ITS (internal transcribed spacer) region and DNA extraction [51], the technique allows for differentiation of the Babesia spp. [13] with high specificity and sensitivity [49]. Additionally, real-time PCR increases the detection limit considerably [14,52]. In addition to PCR, DNA sequency and DNA microarray systems have shown promising preliminary results with infected bovines [53].
The immunological diagnosis of babesiosis is normally established by indirect immunoflourescence (IFA). This is considered the standard assay for detection of Babesia spp. antibodies; however, other techniques such as Imunoblot, immunochromatography, and ELISA (enzymelinked immunosorbent assay) are also used [49]. Specifically, in IFA when using B. microti as antigen, antibody detection reaches 88-96% in infected patients. The IFA slides containing the antigen are prepared using parasited erythrocytes produced in hamsters [9].
Laboratory alterations include hemolytic anemia with increased reticulocytes, and decreased haptoglobulin [13], followed by increased blood sedimentation rate, thrombocytopenia, leucopenia (with lymphopenia), proteinuria, hemoglobinuria, and elevated liver enzymes (aminotransferases, alkaline phosphatase, and lactate dehydrogenase) [13,54]. Specificallly, the infections caused by B. microti are accompanied by decreased hematocrit and hemoglobin counts, elevated total bilirrubins, reduced haptoglobin, and increased reticulocytes, more consistent with hemolytic anemia. Infections by B. divergens show mainly reduced hemoglobin levels (4 to 8 g/dL), indicative of intense hemolysis [13].
Treatment of babesiosis – in symptomatic patients – is carried out with antimicrobial therapy, after diagnostic confirmation [35]. The most commonly used antibiotics are: (1) atovaquone, azithromycin, and a combination of the two; and (2) clindamycin, quinine, and a combination of the two. The binomial dose/duration of administration of these agents generally varies between adult and pediatric patients [14].
In moderate cases of babesiosis caused by B. divergens and B. venatorum, the use of clindamycin for seven to ten days is widely adopted; in infections caused by B. microti the combination of atovaquone and azithromycin is indicated, for the same treatment period [20]. These antibiotics and their combinations are generally administered orally. The most cited side effects are diarrhea and exanthema (azithromycin and atovaquone). It should be highlighted that due to problems attributed to resistance to the principal antimicrobial regimens, the combinations of quinine and clindamycin or atovaquone and azithromycin have been used [12]. However, while treatment with these combinations is effective, there are reports of problems with parasite persistence and response time in endemic areas. Although resistance to these drugs is not suspected in these cases, they do highlight the need for ongoing monitoring of parasitemia during patient treatment [20].
Asymptomatic patients infected with B. microti do not require treatment, unless species of Babesia spp. are detected by blood smear or PCR over more than three months. Symptomatic patients should not be treated if the blood smear or PCR are negative. Treatment is only recommended in case of pathogen detection [13, 25].
In the United States, patients without risk factors or baseline disease usually present low parasitemia, and recovery in most cases without need of specific chemotherapy [37]. More severely affected patients are generally treated with clindamycin 300-600 mg four times daily (by intravenous route), associated with quinine 650 mg thrice daily (by oral route) for seven to ten days. Moreover, some critical patients may require supportive treatment, including: antipyretics, vasopressors, blood transfusion and mechanical ventilation [9].
Chemotherapy and blood transfusions are recommended in Europe and in the more severe cases in the US. The objective of these procedures is to ameliorate parasitemia levels [13,55].
The best preventive measure for babesiosis is avoiding exposure to the habitats of the tick disease vectors, especially in endemic areas and between the months of May and September [13]. Specifically in the case of Ixodes scapularis, they are most commonly found in areas with shrubs and grasses according to the cycle of this vector [9], and activities in such locations require attention.
To date, no vaccine is available, however, efforts have been made to produce one. Despite this, simple efforts help in preventing tick bites and the infections they transmit [9].
In situations of absolute need to visit exposure risk areas, long-sleeves, pants, and socks should be worn, and insect repellents used, especially those containing permetrin (on the clothes and DEET (diethyltoluamide) on the skin [14]. Wearing light colored clothing facilitates visualization and subsequent removal of the ticks [9]. In these areas, care should be taken to walk on open trails, remaining in the center of the path and avoiding exposure to grasses where there is a higher concentration of ticks [9].
Reduction of transmission via blood transfusion can be achieved by exclusion of donors from endemic areas or those with a recent history of tick bites (within two months of donation) [56]. The possibility of babesiosis should always be remembered upon differential diagnosis of transfusion transmitted infections.
Final Considerations
The emergence of babesiosis together with the cosmopolitan distribution of ticks and the environmental changes that favor their cycle are factors that have elevated the zoonotic potential of the disease. New species of Babesia spp. have been described in distinct geographical regions and demonstrate the potential for transmission to humans [57]. The increased diversity and incidence of infections over the recent years has been attributed to better diagnosis, greater awareness on the part of the public, greater contact with natural areas and consequently with the vectors, as well as to environmental changes and the increased number of immunosuppressed individuals [15]. Autoimmune hemolytic anemia (AIHA) may be triggered by babesiosis [58].
Distributed worldwide, little is known about the prevalence of Babesia in countries where malaria is endemic, because of the difficulty in distinguishing it from Plasmodium. There are differences in the species that cause babesiosis, in relation to the regions where the disease occurs. B. divergens is the cause of most cases on the European continent and in the US, B. microti is the principal cause, especially in the northeast and Midwest regions [9].
In Brazil the cases are more restricted, however, another form of transmission – which occurs among travelers – merits attention, given the increased number of travels over the last decades. In this context, information about the transmissible illnesses at the destination is important. Specifically, prevention of babesiosis requires care in endemic areas. In terms of patient care, it is also important to carefully identify the causes of the disease presented, including knowledge of the natural history and place of travel, in order to precisely carry out anamnesis and physical exam [59,60]. Thus it is imperative to know the natural history of the vectors and the species of Babesia that infect humans [15].
Thus, care with education and training of health professionals in relation to this zoonosis is becoming essential, and is fundamentally important in recognizing the disease. Outbreaks may occur as a result of the pathogen’s adaptability to its hosts and climate, which also highlights the importance of the measures mentioned above. Familiarity with the symptoms triggered by babesiosis together with knowledge of efficient diagnostic methods for confirmation is fundamental to establish early treatment and the necessary prophylactic measures.
Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) e CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico).
Download Provisional PDF Here
Article Type: Review Article
Citation: Miguel PSB, Gomes AP, Nogueira CI, Barbosa AS, Bevilacqua PD, Santana LA, et al. (2017) Human Babesiosis and Travelers’ Health: Clinical and Public Health Issues. J Epidemiol Public Health Rev Vol 2(4): doi http:// dx.doi.org/10.16966/2471-8211.150
Copyright: © 2017 Miguel PSB, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access