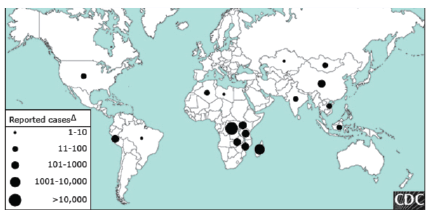
Figure 1: Global Distribution of plague
Global distribution of plague, reported cases by country between 2000
and 2009.
Reproduced from: Center for Disease control and prevention, Division
of vector-borne Diseases.

Luiz Alberto Santana1 Savio Silva Santos2 Jorge Luiz Dutra Gazineo3 Andreia Patricia Gomes1 Paulo Sergio Balbino Miguel1 Mauro Geller2,4 Rodrigo Siqueira-Batista5,6,7*
1Laboratory of Pathogens, Department of Medicine and Nursing, Federal University of Viçosa, Viçosa, MG, Brazil*Corresponding author: Rodrigo Siqueira-Batista, Department of Medicine and Nursing, Federal University of Viçosa, MG, Brazil, Tel: (31) 3899-3978; E-mail: lmecs@ufv.br
The bacteria Yersinia pestis is the etiologic agent of plague, one of the most devastating diseases in the history of humanity. The zoonosis - transmitted by infected fleas - is severe and distributed among countries of Africa, the Americas, and Asia. Humans are highly susceptible to the disease, and maybe infected directly or indirectly. Plague may reappear after decades of silence - with epidemic potential - which can aggravate its public health impact in the various regions. Additionally, due to its easy transmission and dissemination, plague is included with anthrax, botulism, smallpox, tularemia, and viral hemorrhagic fevers (Ebola, Marburg, and Arena virus) in Group A of potential bioterrorism agents. This article presents the main clinical-epidemiological elements of the plague, emphasizing its etiology, transmission, pathogenesis, clinical aspects, diagnosis, treatment, prevention, and aspects relating to bioterrorism.
Plague; Yersinia pestis; Bioterrorism; Xenopsylla cheopis; Zoonosis
The plague is a systemic zoonosis caused by Yersinia pestis, a potentially severe disease that has great historical and clinical-epidemiological significance. Distributed in Africa, Asia and in the Americas, transmission to humans arises from the bite of an infected flea - usually of the genus Xenopsylla - or direct contact with infected mammals [1]. In Latin America from 1899 to 2012, plague has been present in at least one year in 14 of the 25 countries making up the region, with foci remaining in six countries: Argentina, Venezuela, Bolivia, Brazil, Ecuador, and Peru. In Brazil, specifically, between 1980 and 2012, evidence of plague was found in six states (Bahia, Ceara, Minas Gerais, Paraíba, Pernambuco, Rio Grande do Sul) [2]. Yersinia pestis is a gram-negative and highly infectious bacteria [3] and recent DNA studies have shown persistence in the environment over the centuries [4].
Historically, the etiologic agent of the plague has been associated with three large pandemics. The first occurred during the reign of the Roman Emperor Justinian, in the sixth century, resulting in millions of deaths [5]. The second, known as “black death” or the black plague, began around 1347, persisting in Europe until the beginning of the nineteenth century [6] and decimated approximately one third of the European population. The third, arising from mainland China occurred from 1895 to 1930, and caused the death of approximately 12 million people, especially in India [5].
Throughout the years, on all continents, Y. pestis has been transported by rats on ships to port cities. In the Americas and in Africa, infection spread among the various native rodent species, making eradication very difficult [1]. Currently, although the number of notified cases of plague is relatively low, the disease still poses a threat to humanity due to its rapid propagation and clinical evolution, with high lethality rates if not treated early [1]. Additionally, in the United States in 2015, six states registered 11 cases of human plague, four of them in Colorado, the majority of which were among male patients [7]. Although the plague does not compare in numbers to cases of malaria, AIDS, and tuberculosis, it should not be ignored, given the high pathogenicity, rapid propagation [1], and persistence of the pathogen in the environment [4], enabling it to emerge or re-emerge after decades of silence [2].
The objective of this paper is to present the principal aspects of plague and etiological agent that cause plague and the clinical and therapeutic implications of its interaction with the human host.
The genus Yersinia includes 15 species, with three (Yersinia pestis, Yersinia enterocolitica, and Yersinia pseudotuberculosis) considered important human pathogens [8]. The first registered isolation of Y. pestis dates from 1894, in Hong Kong by Alexandre Yersin, from an infectious outbreak spread from mainland China [9]. The bacterium is cosmopolitan in distribution, and is generally isolated from rodents and the fleas that parasite them. It is a Gram-negative microorganism; rod shaped coccobaccilus, non-spore forming and immobile, non-lactose fermenting, urease and indole negative, and included in the class Gammaproteobacteria and the family Enteroacteriaceae. Its optimum temperature for growth over 48 hours in MacConkey or blood agar is 28°C [10].
Isolation of Y. pestis is usually performed in distinct culture medicums such as brain heart infusion agar (BHIA), sheep blood agar, and MacConkey agar [11]. However, the BIN medium is considered the most efficient for isolation and recovery of Y. pestis. The final composition of BIN is formed by infusion of brain and heart agar, with addition of selective agents irgasan, cholate salts, crystal violet and nystatin [12].
Y. pestis is genetically close to Yersinia pseudotuberculosis (97% identical nucleotides) [13]. Genetic sequencing studies and molecular phylogeny suggest that Y. pestis is a recently evolved clone of Yersinia pseudotuberculosis [14]. However, the infection patterns are distinct between these species, and the disease caused by Y. pestis is fatal in the absence of treatment, while Y. pseudotuberculosis is, in most cases responsible for triggering nonfatal intestinal manifestations [13].
Adaptation of the bacterium to fleas is attributed to acquisition of plasmids [15], modification of the LPS (lipopolyssaccharide) component on the external bacterial membrane [16], and the loss of genes that enabled survival in the mammalian intestinal tract [15]. Specifically, survival of Y. pestis in the median intestine of fleas is dependent on the enzyme phospholipase D, coded by the Ymt gene, responsible for protecting the pathogen from bacteriolytic agents present in this region [17]. The persistence of Y. pestis in the flea intestinal tract depends on the ability of the bacteria to form multicellular aggregates too large to pass to the feces [18]. Y. pestis is described in four biotypes (Antiqua, Medievalis, Orientalis, and Microtus), subdivided according to glycerol fermentation capacity and conversion of nitrate to nitrite. The recent genome analysis of a strain of Y. pestis associated with the “Black Plague” indicated that it was more closely related to the Orientalis biotype than with the Medievalis [15]. This interaction between Y. pestis and the transmitting flea results in colonization and successful transmission, considered to be a result of adaptive evolution requiring few genetic alterations. Among these alterations are the loss of insecticide activity, increased resistance to antimicrobials and capacity to form biofilms demonstrated by Yersinia in the interior of the vector [18].
Several species of flea are ectoparasites to humans and can, in this interaction, transmit diseases [19]. Although more than 80 species of fleas have the potential to transmit plague by Y. pestis, Xenopsylla cheopis (Siphonaptera: Pulicidae), popularly known as the Oriental rat flea, is the most efficient primary vector [20]. Another recently recognized vector is Synopsyllus fonquerniei (Siphonaptera, pulicidae) [21]. Other disease vector include Ctenophtalmus bacopus, Ctenophtalmuscabirus, Dinopsyllus lypusus, Pulex irritans and Xenopsylla brasiliensis [22]. The risk of human infection is increased in epizootic periods, which are generally associated with elevated rodent and flea densities [20].
Plague is a zoonosis of domestic animals (dogs and cats), nonhuman primates, insectivorous mammals (shrews and porcupines), predators (foxes, possums, and beavers), camels, lagomorphs, and especially wild rodents [23]. Humans are considered accidental hosts since they present no contribution to the disease cycle. Disease transmission occurs most often by flea bites, especially those of the species Xenopsylla cheopis and Xenopsylla brasiliensis. However, there are other forms of transmission, such as: (1) The scratch or bite of an infected animal; (2) Inhalation of particles eliminated by animals carrying respiratory Y. pestis infection, or; (3) During autopsy of infected animals [24]. Handling of the pathogen by laboratory professionals, without use of appropriate biosafety equipment, is another form of transmission that should be mentioned [25].
Plague is principally distributed in countries of the African, American, and Asian continents (Figure 1). Between 2000 and 2009, the number of worldwide registrations of the disease was approximately 21,725, with 1,612 deaths in 16 countries. The number of cases officially notified to the World Health Organization (WHO) is likely to be considerably smaller than the real number, as countries in endemic areas do not register all cases and there are difficulties in diagnosis of the disease in these countries [26].
Figure 1: Global Distribution of plague
Global distribution of plague, reported cases by country between 2000
and 2009.
Reproduced from: Center for Disease control and prevention, Division
of vector-borne Diseases.
In Africa, plague has probably been present since before the third pandemic and hundreds of thousands of human infections have been reported in 26 countries since 1877 [27]. Over the last few years, roughly 90% of worldwide cases of plague have been reported in Africa. Among these the most recent occurred in Argelia, in 2003, in Libya in 2009, probably resulting from reactivation of Y. pestis in foci thought to be extinct (Argelia) or dormant (Libya) [28]. In Madagascar, an outbreak was also reported in 2014, with 482 cases notified. In this country, plague occurs primarily as a rural disease, however, an urban epidemic has been described (in the coastal city of Mahajanga), an outbreak of pneumonic plague in the northeast region in 2011, and there is also sylvatic transmission involving rodents and fleas endemic to the region [18,29,30].
In the US, 1,006 cases of human plague were notified between 1900 and 2012. The median age of affected patients was 29 years (varying from <1 to 94 years), and 644 (65%) of the 992 patients whose gender was reported were male [31]. Currently, in Latin America, human cases and natural foci of the disease are present in Bolivia, Brazil, Ecuador, and Peru [2]. In Brazil, 3,693 cases were reported between 1958 and 2008, and currently there are two independent focus areas: the Northeast, including an extensive area corresponding to the Polígono das Secas (drought polygon) that extends from the state of Ceará to the north of Minas Gerais, and the Serra dos Órgãos (in the state of Rio de Janeiro), within the municipality limits of Nova Friburgo, Sumidouro, and Teresópolis [23].
Transmission and distribution of the plague is modulated by environmental and seasonal conditions. In the United States and in Asia, epidemics occurred at the end of winter, a time when rodents typically leave their dens after hibernation. However, in other areas the dynamic of the fleas is different, given the vector’s sensitivity to temperature and air humidity [18,32]. In fact, low temperatures delay bacterial proliferation [18]. It is therefore difficult to explain the dissemination of the disease solely by rat fleas in some pandemics, especially in the winter, a time of lesser activity for many vector species. In this sense, transmission by droplets - in close contact with the sick - should be highlighted as being related to pneumonic plague; nevertheless this form of transmission alone is not enough to explain some pandemics. With regards to the second plague pandemic - that devastated Europe from 1347 to the early nineteenth century, one study provided scientific evidence explaining how the climate caused repeated reintroductions of the bacteria in European ports, from disease reservoirs in Asia, with a mean interval of 15 years (variation of 14 to 16 years) [6]. Thus, climate and meteorological phenomena (such as El Niño) appear to influence all three components (i.e. bacteria, vectors, and hosts) in the transmission chain and may explain some of the variability of the disease from small and regional to large-scale outbreaks [33-39].
The role of the soil as a reservoir for Y. pestis remains controversial, as it is proposed that Y. pestis may survive intracellularly to the protozoa in this environment. Additionally, transmission of plague by Pediculus humanus humanus requires confirmation, since there is no evidence in the literature consulted that human ectoparasites naturally transmit the disease [40].
Flea infection occurs during feeding from a host with Y. pestis bacteremia, pathogens that colonize, multiply, and block the interior of the arthropod intestines. When feeding, fleas regurgitate bacteria in the bite wound [41], and the bacteria are introduced in the capillary system of the dermis [42]. At this time, most of the bacteria inoculated express the F1 antigen; subsequently, the pathogens undergo phagocytosis by polymorphonuclear leukocytes [43]. However, some bacteria escape phagocytosis, and due to lack of phagocytic activity during inoculation, are captured and transported by the macrophages, reaching the nearby lymph nodes. Consequently, there is stage change from intracellular multiplication to intense extracellular multiplication [41]. Bacterial dissemination in the bloodstream may result in primary septicemic plague, and, if the infection persists, dissemination of bacteria may - via the circulatory system - reach the spleen, liver, and lungs. In these organs, in case of extensive bacterial replication, it is possible for host death by septic shock and multiple organ failure to occur [42].
During replication in the lymph nodes and surrounding regions, Y. pestis produces local inflammation (bubo). Subsequently there is expression of the bacterial antiphagocytic system, resulting in decreased capacity of the host response, which likely facilitates peripheral dissemination and recovery by a vector that interacts with the host [43]. Microscopic exam reveals normal lymph node architecture, and also shows presence of polymorphonuclear cells, hemorrhagic necrosis, and extracellular presence of the bacilli [44]. In this phase, bacteremia is very common and sepsis may occur, with hemorrhagic and purulent lesions on multiple organs [45]. Resulting from sepsis, there is also often occurrence of disseminated intravascular coagulation [7], leading to circulatory shock and multiple organ system failure [42]. Symptoms include acute onset of fever, chills, prostration, abdominal pain, nausea, and vomiting. Eventually, purpuric lesions may arise on the skin, and there may be a need to amputate toes and fingers [7].
Another form of plague transmission by Y. pestis arises from direct inhalation of aerosol by the lungs, which results in primary pneumonic plague [42].
The plague presents three more common and distinct clinical forms: the bubonic form (bubonic plague), septicemic form (black plague) and the pulmonary form [46].
The most frequent form is bubonic plague, representing between 80 to 95% of plague cases, and arising between two and ten days after Y. pestis innoculation [46]. This form presents as painful adenitis near the site of the flea bite (bubo), most often on the inguinal-crural region. Additionally, it may affect the cervical and axillary lymphatic chains, especially in children. Aside from the bubo, high and continuous fever may arise, accompanied by chills, fatigue, and headache. Delirium, prostration, shock, and death may occur; however most cases evolve benignly. The swollen lymph nodes are oval, extremely painful, and often vary between one and ten centimeters in diameter, and the surrounding skin may be hot and erythomatous. Palpation is painful, with a feeling of an irregular, oval mass, due to the junction of multiple lymph nodes. Patients generally seek medical assistance in the later phase of the disease, and in this phase there is often lethargy, prostration, agitation and even delirium upon physical exam. Fever may occur between 38.5 to 40.0°C. Tachycardia is frequent, and systemic arterial hypotension is not uncommon. There is palpable hepatosplenomegaly, with both organs presenting as soft and painful to palpation. The fatality rate of bubonic plague is 50 to 60% of untreated cases, and less than 5% with use of appropriate antibiotic therapy [47,48].
The second form is septicemic plague, which generally evolves from the untreated bubonic form; however, bacterial proliferation sometimes occurs in the organism in the absence of the bubo. The disease affects various organs and systems, and the most common signs are tachycardia, systemic arterial hypotension, sub-mucosal hemorrhages, echymoses, and vasculites accompanied by necrotic lesions in the fingers and skin. Death of the affected patient may occur within 24 or 48 hours. Diagnosis in the absence of the bubo may be difficult, since the clinical picture is similar to sepsis of other etiologies [49,50]. Roughly one-third of these patients perish before diagnosis of plague is established. The lethality rate of the cases of untreated septicemic plague is close to 100% [47].
Pneumonic plague presents two forms, primary and secondary. Transmission generally occurs among people by respiratory droplets [10,47]. The secondary form is more common and evolves from the bubo by hematogenous or another as yet unidentified pathway. An estimated 10% of patients diagnosed with the bubonic form, with adequate late stage treatment, evolve to the pneumonic form. The primary form of the disease, in turn, occurs by direct inhalation of the bacteria through contact with patients, infected animals, or by occupational exposure in laboratories [5,24,51,52]. The pneumonic form is usually a severe infection – with rapid evolution and is often accompanied by sepsis. Patients in this condition may present productive cough with purulent expectoration, sometimes with bloody sputum or even hemoptysis. This is followed by thoracic pain, dyspnea, tachypnea, and systemic inflammatory response syndrome [49,50]. Chest radiograph may show consolidations, bronchopneumonia or cavitations. Although a rare clinical presentation (approximately 2% of notified cases) primary pneumonic plague has a high lethality rate (greater than or equal to 93%) when untreated [51].
The bacterimia arising from any form of plague may, in rare instances, lead to meningitis by Y. pestis. The clinical and laboratory findings are similar to those described in other bacterial meningitis; rarely, pleuritis, endophthalmitis, and myocarditis by the etiologic agent may also occur [53].
Clinical diagnosis of plague is established by patient history and by clinical exam. In this sense, immediate attention should be given to patients presenting: (1) Fever with lymphadenomegaly after contact with dead rodents, in a region of occurrence for this zoonosis; (2) Occurrence of lymphadenomegaly with fever and systemic arterial hypotension, not explicable by other causes; and (3) Occurrence of pneumonia with bloody expectoration and presence of Gram-negative cocobacilli in the sputum [11]. Therapy should be initiated upon diagnosis of gramnegative bacilli or cocobacilli in lymph node aspirate, due to the rare occurrence of secondary lymphadenomegaly in diseases caused by other bacteria. Another suggestion is the identification of the Gram-negative bacilli in sputum of patients with community acquired pneumonia, in endemic areas, especially when accompanied by leukocytosis with thrombocytopenia, which makes the clinical picture very suggestive of plague [7].
The materials obtained from the patient (aspirate of a floating lymph node bubo or sputum obtained in the pulmonary form or peripheral blood smear) show by Gram, Geimsa or Wright staining - on microscopy - the classical bipolar morphology of Y. pestis [7]. Additionally, microbiological cultures of material suspected of infection and blood (culture) are performed by culture in blood or Mac-Conkey agar, although the growth of Yersinia is relatively slow [11].
Serological confirmation in the acute phase of the disease requires a fourfold increase in antibody titers against the F1 antigen of Y. pestis. The use of the passive hemagglutination test may be useful, given that a single title above 1:16 is suggestive of the diagnosis. Polymerase chain reaction (PCR) is very useful in the diagnosis of plague, especially in view of bioterrorism, given the speed, sensitivity, specificity, and ease of performance [54].
The drug of choice for the treatment of plague is streptomycin, at the dose of 15 mg/kg (maximum dose of one gram), at 12 hour intervals, by intramuscular route (IM) for 10 days [10,47,55,56]. Another alternative - used when streptomycin is unavailable - is gentamycin, at a dose of 5 mg/kg/day, or 2 mg/kg as an initial dose followed by 1.7 mg/kg every 8 hours, by IM or intravenous (IV) route, for 10 days [10,47,48]. A retrospective analysis of 50 plague cases diagnosed in New Mexico (USA), between 1985 and 1999, suggests that gentamycin administered alone or in combination with doxycycline is at least as effective as streptomycin. The 36 patients treated with gentamycin survived without complications [57]. The randomized clinical trial of 65 patients with plague, performed in Tanzania, showed that 94% of subjects treated with gentamycin had high rates of favorable clinical response and low rates of associated side effects [58]. Gentamycin is generally considered safer for administration to pregnant woman and children, as compared to streptomycin [48].
Patients intolerant to aminoglycosides are usually treated with tetracycline, with an initial dose of two grams by oral route (VO), followed by two grams per day divided in four ingestions. Another alternative is doxycycline, at the initial dose of 200mg, at 12 hour intervals, VO, on the first day, followed by 100mg, VO, twice daily. Care should also be taken with patients exhibiting central nervous system involvement (Y. pestis meningitis). In these cases, chloramphenicol is used due to its greater capacity to cross the blood brain barrier; in this case, the initial dose is between 25 to 30 mg/kg. The quinolones - such as levofloxacin (500mg/ day), ciprofloxacin and moxifloxacin (400mg/day) - have been previously used in animal studies and in-vitro pharmacodynamic models [59]. However, clinical data for the use of these drugs to treat plague in humans is lacking. It is important to note that levofloxacin was approved by the FDA in 2012 for treatment and prophylaxis of plague.
The medical literature shows that only two people diagnosed with plague presented pathogens resistant to streptomycin. The cases occurred in Madagascar, in 1995, and survival of these patients was attributed to concomitant use of sulfamethoxazole-trimethoprim (together with streptomycin). The cases were caused by strains of Y. pestis that possessed resistance plasmids (IncA/C) transferred from other bacteria [48]. However, these resistant strains do not persist in the environment, since all subsequent cases of plague have been caused by strains of bacteria susceptible to the antibiotics tested [60]. In fact, one analysis of the antibiogram of 392 clinical samples of Y. pestis from 17 countries in the Americas, Africa, and Asia, did not find any resistance to eight of the antibiotics used for plague treatment or prophylaxis [61].
In more severe cases, in addition to specific antibiotic therapy, supportive treatment should also be instituted (dialysis, mechanical ventilation, vasoactive amines, blood transfusion, among others), as needed [47,48]. There is no evidence the use of glucocorticoids is of benefit in the adjuvant treatment of plague [48].
Since the beginning of the XX century, two types of vaccine against plague have been available for use in humans: a whole-cell inactivated vaccine and a live attenuated vaccine. None of these immunobiologicals is currently available in the US, despite the live attenuated vaccine being found in China and countries of the old Soviet Union. Nevertheless, fear of a possible bioterrorist attack stimulated research of new immunization strategies for plague [47,48]. Recombinant subunit vaccines based on the RF1 and RV antigens are the most promising perspectives and are being studied in clinical trials (Phase I and II), prior to the licensing process. New live attenuated anti-Y.pestis vaccines should trigger immune responses - humoral and cellular - against a variety of relevant antigens, providing a greater protection against all forms of the disease, including pneumonic plague [62,63].
Post-exposure prophylaxis should be initiated up to seven days after exposure to risk for: a) Individuals who have had intimate contact with confirmed or suspected pneumonic plague patients; b) Family members; c) Caregivers; d) Laboratory workers exposed to an accident that may have generated infectious aerosols [48]. The prophylactic antibiotics of choice are doxycycline (at the adult dose of 100mg, VO, every 12 hours) or ciprofloxacin (500mg VO every 12 hours) or levofloxacin (500mg VO every 24 hours). This prophylaxis should be maintained for seven days [10,47,48,55]. Chloramphenicol (25 mg/kg, VO, every 6 hours) would be the alternative drug [47]. The WHO also recommends, for post-exposure prophylaxis: tetracycline (one to two grams VO divided in intervals of six or 12 hours) or sulfamethoxazole-trimetoprim (1.6 grams VO divided in intervals of 12 hours) [64]. Pre-exposure prophylaxis may be initiated for people travelling to areas with active plague transmission, during a short time interval, in circumstances where there is an inevitable risk of acquiring the disease (presence of large quantities of fleas or cases of pneumonic plague) [47].
Overall, plague requires only standard precaution measures, but patients with pneumonic plague, both primary and secondary, require droplet precautions [10,47]. Isolation of patients with pneumonic plague (suspected or confirmed) should be maintained for at least 48 hours after initiation of antibiotic treatment or until clinical improvement [47,65]. In Brazil, all suspected and confirmed cases of plague should be notified to the hospital epidemiology service or the hospital infection control commission (in the absence of the former) and also to the municipal and/ or state Health Department [47]. Microbiology laboratory workers should be alerted to any clinical sample suspected of harboring Y. pestis, because they are required to adopt Biosafety Level-2 precautionary measures for general procedures and Biosafety Level-3 for procedures that may produce aerosols or droplets [47]. In fact, in Brazil, two professionals (one who dealt with rodents in the field and another who worked in the National Plague Reference Laboratory) possessed serum antibodies for Y. pestis, but had no previous diagnosis of the disease. This information is relevant to minimize the occupational risk and evaluate the efficacy of current biosafety practices in the handling of this etiologic agent [66]. Given that Y. pestis is very sensitive to sunlight and heat, it does not usually survive long outside of the host. Environmental decontamination is performed using a 0.5% hypochlorite solution or bleach diluted at 1:10 [47].
The best preventive measure is to avoid exposure to risk situations for the disease. Recommendations include: handling rodent and other wild animal carcasses with gloves; avoiding areas where a high lethality of commensal rats has been observed; use of gloves when manipulating potentially infected animals; control of peridomestic rodents; flea control for pets with appropriate insecticides; use of appropriate insect repellents on the skin, clothes, shoes, and camping equipment; use of long pants; cooking of meat at temperatures above 71°C [51,67]. A study performed in Uganda corroborates that, in addition to the existing preventive measures, permanent efforts should be made for ongoing education and awareness on the part of health professionals about the clinical picture of the disease and the importance of immediate and adequate antibiotic therapy [19].
Plague has been used as a biological weapon since the XIV century. Later, during the Second World War, the Japanese army supposedly launched clay pots containing fleas infected with Y. pestis in populated areas in China, on several occasions, resulting in small plague outbreaks in at least three cities. During the Cold War, The United States and the former Soviet Union were involved in the development of biological weapons using aerosolized Y. pestis, prior to the international convention of 1972, prohibiting production, storage, and use of biological weapons [47,48].
Plague, together with anthrax, botulism, smallpox, tularemia, and the viral hemorrhagic fevers (Ebola, Marburg, and Arenavirus) are in Group A of the CDC list of high priority potential bioterrorism agents, since they can: (1) Be easily disseminated or transmitted from person to person; (2) Result in high lethality and have the potential to cause significant public health impact; (3) May cause public panic and social disruption; (4) Requires special action for public health preparation [68].
Pneumonic plague would be a devastating condition in the context of biological warfare [55]. According to a WHO estimate, if 50 kg of Y. pestis were aerosolized and spread over a city of five million people, pneumonic plague could affect up to 150,000 people and result in 36,000 deaths. Y. pestis would remain viable in the air for one hour and would travel up to 10km in the direction of the wind [47]. However, a recent mathematical simulation model suggests that risk of secondary disease transmission is very low for most people [69]. Additionally, Y. pestis does not form spores and does not survive well outside animal bodies (humans and nonhumans). For these reasons, no biological weapon using aerosolized Y. pestis has been successful to date [46].
In a deliberate bioterrorist attack, primary pneumonic plague - in place of secondary propagation of the bubonic or septicemic form - would occur one to three days after inhalation of the bacteria released or after transmission of droplets from an infected individual. The initial presentation of pneumonic plague is nonspecific and difficult to differentiate from other community acquired pneumonias in its early stages [70]. Hemoptysis, a unique characteristic, present in more advanced stages of the disease (from one to several hours before death) [5] and rapid progression to respiratory failure and death occur more often in plague than in community-acquired pneumonias caused by other etiologic agents [70].
In a clinical-epidemiological context in which a small number of patients with plague require treatment, use of streptomycin or gentamycin is recommended, by parenteral route [10]. However, in a context in which there is a large number of cases (mass casualty setting), intravenous or intramuscular antibiotic therapy may not be possible for logistical and operational reasons, thus, in this scenario oral antibiotic therapy is recommended, preferably with doxycycline 100mg VO every 12 hours or ciprofloxacin 500 mg VO every 12 hours or levofloxacin 500 mg VO every 24 hours [10,48]. Plague vaccines are not currently available in the United States, and due to the short incubation period of the disease, they would not have an important role in an immediate response to bioterrorism [48].
Due to its high epidemic potential, plague may negatively impact public health, tourism, and international commerce. The disease is usually distributed among the nations of Latin America, especially Bolivia, Brazil, Ecuador, and Peru [2]. Other endemic áreas are added to this list, in countries in Africa and Asia [26].
The success of the interaction between Y. pestis and rat fleas allows transmission cycles that resulted in profound historical consequences, manifested by pandemics in humans. This route of transmission is considered an ecological change, demonstrated by Yersinia pseudotuberculosis, the species from which Y. pestis derived [18]. Additionally, the disease may reemerge after decades silence, and may further aggravate the public health impact and the epidemic potential [2]. The potential for re-emergence of plague after years of silence demonstrates the importance that should be given to patient care and education of health professionals. Of equal importance is familiarity with the symptoms of plague, and the knowledge of the confirmative diagnostic methods.
The main risk factors for infection are considered to be the behavioral ones that maximize direct or indirect contact with rodents and their parasite fleas. Essential to adequate and early treatment is clinical suspicion, and recommended treatment is with aminoglycosides and tetracyclines [31]. However, the fluoroquinolones are also considered effective.
Download Provisional PDF Here
Article Type: Review Article
Citation: Santana LA, Santos SS, Gazineo JLD, Gomes AP, Miguel PSB, et al. (2016) Plague: A New Old Disease. J Epidemiol Public Health Rev 1(4): doi http://dx.doi.org/10.16966/2471-8211.128
Copyright: © 2016 Santana LA, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access