Abstract
The adrenal cortex and medulla of mammals have complex anatomical and functional relationship. Anatomically: the cortex and medulla are connected by adrenal portal vascular system thereby the medulla is irrigated by cortisol-rich blood; the cortical and medullary cells are intermingled within the normal adrenal gland. Functionally: adrenaline synthesis by the enzyme phenylethanolamine N-methyltransferase requires high cortisol concentrations. Corticosteroids are reportedly produced from precursors of cortisol in extra-adrenal pheochromocytoma tissue in vitro.
Interactions of the cortex and the medulla may shed light on some symptoms of patients with Adrenal Insufficiency (AI), which cannot be explained by the failure of either cortex or medulla alone. Following examples are retrieved from the author’s clinical files as well as from literature: persistent fatigue of a few patients with AI despite being appropriately supplemented with hydrocortisone; absence of autonomic nervous system symptoms in AI patients during hypoglycemic episodes; elevated aldosterone levels of an adult patient with salt-losing 21-hydroxylase deficiency in neonatal period; normal or elevated cortisol levels in critically ill patients with sepsis syndrome notwithstanding suppressed hypothalamo-hypophseal system. These observations are possibly explained by impaired adrenaline production due to insufficient cortisol supply to the medulla or by interactions of cortical and medullary cells within the adrenal gland.
The functional relation between the cortex and the medulla may be un-noticed in man with the intact adrenal gland. The understanding of the cortico-medullary collaboration in immediate and sustained adaptation to altered homeostasis in man would be advanced after accumulation of more clinical cases of adrenal dysfunction.
Keywords
Adrenaline; Adrenal portal system; phenylethanolamine N-methyltransferase; Persistent fatigue
Abbreviations
TH-Tyrosine Hydroxylase; AAD-Aromatic L-Amino Acid Decarboxylase; DBH-Dopamine β-Hydroxylase; PNMT-Phenylethanolamine N-Methyltransferase
Background
The anatomical relation of the adrenal cortex and the adrenal medulla has been well known. Both the cortex and the medulla are located above the kidneys inside the common capsule. The medulla is enclosed by and juxtaposed to the cortex. Cortisol has a trophic role in the medulla by the juxtaposition, i.e. cortisol-rich blood is supplied from the cortex to the medulla via adrenal portal vascular system (Cf. section 2), which is required to keep the amount of adrenalinesynthesizing enzyme, Phenylethanolamine N-methyltransferase (PNMT) [1]. Hence, the production of Adrenaline (AD) is tightly dependent on cortisol supplied to the medulla.
Conversely, there are a couple of evidences of medulla’s influence on adrenocortical steroidogenesis: Transformation of pregnenolone to subsequent intermediates in the cortisol synthesis are observed in pheochromocytoma tissue in vitro [2]; medullary cells influence activities of cortical cells in a paracrine manner through intermingling of cortical and medullary cells in the adrenal gland, which is demonstrated by immunohistochemical stain and by electron microscopy [3].
Carballeira and Fishman proposed “the adrenal functional unit” for cortico-medullary interactions in their review of 1964 [2]. In the review of 2011, Bornstein and coworkers recapitulated corticomedullary interactions, i.e. contact of cortical and medullary cells within the adrenal gland, interdependence of these cells on hormone synthesis as well as on responses to stress, and findings of in vitro studies, animal models and clinical studies [4].
In the subsequent sections, the structural and functional relations of the cortex and medulla are reviewed. Clinical manifestations of altered cortico-medullary interactions are presented, which were observed in a few patients with latent Adrenal Insufficiency (AI) and in cases from literature. Herein, latent AI is defined by lack of some of signs and symptoms of Addison’s disease such as hyper pigmentation, weight loss, hypotension, etc., impaired cortisol responses to provocation tests, and amelioration of symptoms after corticosteroid supplementation [5].
The Anatomy of the Adrenal Gland and the Vasculature
The adrenal gland is divided into three parts in pathological examination, i.e. the head, the body and the tail, from the medial to the lateral [Figure 1]. The medulla is unevenly contained in the adrenal gland, i.e. the ratios of cortex/medulla are 4:1. 15:1, respectively and medullary tissues are absent in the tail [6]. The cortex is divided into three layers, i.e. zona glomerulosa, zona fasciolata, and zona reticularis. Three arteries supply the adrenals via secondary branches: phrenic artery, aorta, and renal arteries whereas there are no arteries directly supplying the medulla. Arteries entering the adrenals through the Gerota’s fascia form the first plexus (capsular plexus). The branches from the capsular plexus run through zona glomerulosa, zona fasciolata, and zona reticularis in parallel and form the second plexus (plexus reticularis) in the cortico-medullary border. Branches from the second plexus enter the medulla. Vessels of the cortex and the medulla constitute portal system, which enables supply of cortisolrich blood to the medulla [7] [Figure 1]. This system is contributory to biosynthesis of AD from Noradrenaline (NA) in the medulla as described in the next section.
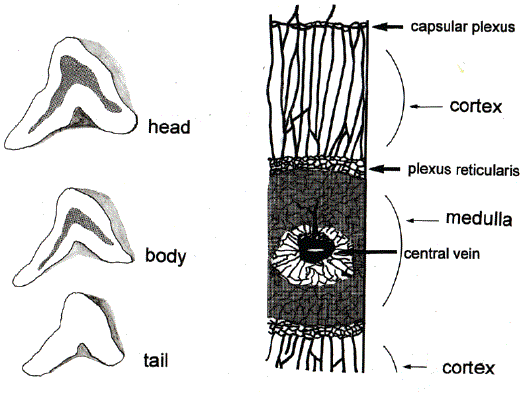
Figure 1: Three Divisions of Adrenal Gland and Vasculature.
Left side, three sections of adrenal gland. The sections of head, body, and tail are shown with the medulla (shaded areas) in the sections of head and body.
Right side, vasculature of head and body, modified from the original by Dobbie JM and Symington T [7].
The medulla of the mammals is enclosed by the cortex. The medullary tissue makes up about 10 % of the weight of the adrenal gland [8]. Human adrenal medulla contains 4.3 times more AD than NA [9]. In cartilage fish, e.g. dogfish, the cortex and the medulla are separated and no AD is contained in the medulla [10].
Biosynthesis of Catecholamines in the Medulla
L-tyrosine is sequentially hydroxylated, decarboxylated, and hydroxylated upto NA in the medulla as well as in the paraganglia. AD is synthesized by methylation of NA by PNMT in the medulla [Figure 2]. Synthesized AD is stored in chromaffin granules of the medullary cells together with NA as well as with many neuropeptides until released into blood by splanchnic nerve stimulation. PNMT exists mostly in the medulla and in small amounts in the brain and the heart. Therefore, plasma AD mainly derives from the medulla [11]. Patients with AI do not produce AD sufficiently because not enough cortisol is supplied to the medulla even they are supplemented with physiological doses of hydrocortisone. On the other hand, AD production is not augmented by excessive cortisol in patients with Cushing’s syndrome [12].
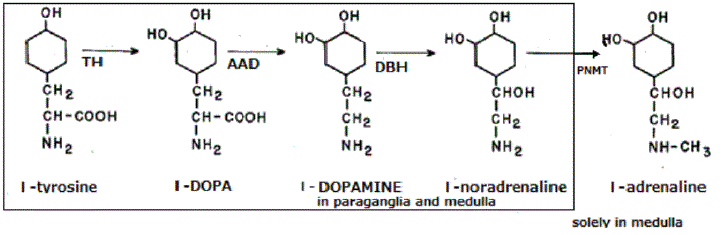
Figure 2: Biosynthesis of Adrenaline in the Adrenal Medulla.
The inner square shows the synthetic processes in the medulla as well as in the paraganglia and the outer square the process in the medulla.
Interactions between Cortical and Medullary Cells
The cortex and the medulla are separated by routine histological examination. Bornstein and coworkers studied cortical and medullary cells of the adrenal gland of experimental animals [13,14] and man [15] by immunohistochemical staining and electron microscopy and found intermingling of cortical and medullary cells in the adrenal gland. Namely, the presence of cortical cells are observed in the medulla as clusters, islets, or single cells by immunostaining with anti17 α-hydroxylase antibodies; rays of chromaffin tissue are revealed in the cortex by immunostaining with chromogranin-A antiserum; the presence of cortical cells in the medulla and of chromaffin cells in the cortex are probed by endoplasmic organelles of respective endocrine organs; the cortical-medullary junction is mainly composed of medullary cells and cortical cells of the zona reticularis. Bornstein’s group continued studies of functional interactions of cortical and medullary cells and postulated an interaction between the cortex and medulla: when medullary cells activated by splanchnic nerve stimulation, medullary cells turn on cortical cells in a paracrine manner by NA and neuropeptides contained in chromogranin granules; the interaction participates in adaptation to severe physical stress as sepsis syndrome [16]. Carbelleira and coworkers reported steroidogenic ability of the medulla as follows: slices and homogenates of pheochromocytoma tissue cannot split cholesterol side chain and transform cholesterol to pregnenolone; the preparations hydroxylase pregnenolone at carbons 21, 17, and 11(C21, C17, C11) with 25 to 40 % capacity of cortical cells; the steroid hydroxylating enzyme activity is detected in the same intra-cellular locations (mitochondria and microsomes) in pheochromocytoma cells; 11-hydroxylation is blocked by metyrapone [2]. Subsequently, they reported synthesis of androstenedione and testosterone from corticosteroid precursors except for cholesterol by homogenates of pheochromocytoma tissue [17].
Studies of AD Production in Patients with AI
Decreased AD production has been reported in adrenal insufficiency by several groups of investigators. Namely, urinary excretion of AD is reduced in patients with idiopathic or tuberculous Addison’s disease [18], those with adrenalectomy [19], and those with secondary adrenal insufficiency [20]. Plasma AD levels of patients with Addison’s disease was lower than those of healthy controls [21]. Plasma AD levels are reduced in corticosteroid-supplemented children with 21OHD as well as in adults having been adrenalectomized bilaterally for either familial pheochromocytoma or Cushing syndrome [22]. In corticosteroid-supplemented patients with glucocorticoid deficiency due to unresponsiveness to ACTH, AD levels were lower in resting state, in upright position, to cold press or test, or following exercise than the levels to these stimuli in control subjects, which was partially compensated by increased NA [23]. A pathological investigation of the adrenals excised from patients with 21OHD revealed deranged formation of the adrenal medulla, i.e. incomplete development of the medulla and extensive intermingling of cortical and medullary cells [22]. Clinical manifestations of decreased AD production were not to referred to in these reports.
Clinical Implications of Impaired AD Production
In the early years of medicinal trials of treatment of AD before corticosteroid treatment, patients were treated with dried whole suprarenal gland by mouth [24] and then with AD, administered either by subcutaneous injection or by rectal instillation [25]. Only small numbers of patients were treated with these agents. Subsequently oral administration of ephedrine, an AD-mimetic, was tried [26]. Results of ephedrine trial were described as follows: “a marked feeling of wellbeing, significant increase in strength, ---Feeling somewhat stronger for several hours ---Feeling of weakness and exhaustion disappeared and the patient felt buoyant, strong and refreshed” [26]. The last quote suggests an association of lack of strength with AD deficiency. The efficacy of ephedrine to alleviate lack of strength, however, has not been studied ever since.
The author has observed the association of fatigue with low AD levels: three patients with latent AI (Cf. background section) had persistent fatigue even after Hydrocortisone (HC) supplementation for two to three months; all symptoms, fatigue as well as other symptoms were ameliorated in other seven patients after HC supplementation in similar periods; The AD levels were less than 0.01 ng/ml in the former and 0.02 to 0.06 ng/ml in the latter. AD levels were measured by chemoluminescence assay after separation by high-performance liquid chromatography. The AD levels of controls were 0.01-0.08 ng/ ml, median 0.03 ng/ml (N=20). Then, these patients continued HC supplementation for more than three months, their complaint of fatigue reduced and their AD levels raised to 0.02 to 0.04 ng/ml (the author, unpublished observation). The resurgence of AD production might have occurred in the three patients by longer duration of HC supplementation. Another example of fatigue and AD is chronic fatigue syndrome (CFS), i.e. patients with CFS have low AD levels in resting state and show attenuated response to exercise [27]. Their cortical function is not referred to in the report.
Hypoglycemia unawareness is considered as a symptom of catecholamine deficiency [28]. Hypoglycemia normally elicits secretion of counter regulatory hormones, i.e. glucagon, adrenaline, growth hormone and cortisol via ACTH secretion and stimulation of Autonomic Nervous System (ANS). When hypoglycemia set in, patients become aware of physical symptoms of warning, i.e. sweating, palpitation, tremor, which are brought in by ANS stimulation together with AD discharge. If insulin-treated diabetic patient has concurrent medullary insufficiency, he would be unaware of hypoglycemia for lack of physical warning symptoms and AD-mediated hepatic glycolysis. One of the author’s patients with AI was treated with insulin for comorbid diabetes mellitus. The patient was troubled by recurrent calf muscle pain without inflammatory signs. The diagnosis of AI was based on 30-minute peak cortisol level of 18.4 μg/dl following 1- μg corticotrophin i.v injection (95% confidence interval of 30-minute peak levels of 40 control subjects, 21.5-23.3 μg/dl) and amelioration of the symptom after HC supplementation. At one time, he was totally asymptomatic in spite of blood glucose of 25 mg/dl. If insulintreated patients have concurrent AI, they have to be warned of neurohypoglycemia without warning ANS symptoms.
Clinical Observations of Medulla’s Influence Over the Cortex
21OHD is an inherited disorder of enzymatic defect in cortisol biosynthesis, characterized by impaired production of corticosteroids, compensatory overproduction of ACTH, and excessive production of precursors of cortisol. Adult patients with 21OHD have obesity, hypertension, and osteoporosis because of high doses of HC to suppress ACTH. Though patients with salt-losing 21OHD need to be supplemented with fludrocortisone in early childhood, adult patients can do without fludrocortisone if ample salt is supplied. The author took care of adult patients with salt-losing such 21-hydroxylase deficiency in neonatal period, who disliked glucocorticoid therapy because of overweight associated with high-dose glucocorticoid to suppress ACTH levels. Without glucocorticoid maintenance therapy, the patient’s cortisol and aldosterone levels were 5-10.6 /dl (reference range, 4.5-21.1) and 32.1-106 (3.6-24.0), respectively [29]. It appears possible that medullary cells of the patient produce cortisol and aldosterone by hydroxylation at C21, C17, C11 from abundant progesterone and 17 alpha-hydroxyprogesterone (precursors of cortisol) even though the medullary cells are unable to split cholesterol side-chain (Cf, section 4).
Critically ill patients with sepsis syndrome have normal or elevated cortisol levels despite suppressed ACTH levels [30-32]. The observations are explained as, stimulation of cortical cells by means of catecholamines and chromogranin granules-derived proteins released by stress-mediated splanchnic nerve stimulation from the intermingling medullary tissue within the cortex [23], elevated free cortisol levels and reduced cortisol-binding protein [32], suppressed cortisol breakdown [31], or participation of endothelin and/or natriuretic hormone [30].
Carballeira and Fishman wrote, “Intimate anatomic coupling of cortex and medulla is likely to have important functional and adaptive correlates [32].” The cortico-medullary collaboration, however, remains un-noticed in daily life of man with the intact cortico-medullary function. A few examples suggestive of altered cortico-medullary collaboration are presented. These observations have to be confirmed in more cases. The understanding of cortico-medullary collaboration in immediate and sustained adaptation to altered homeostasis would be advanced after accumulation of clinical cases of adrenal dysfunction.
Article Information
Article Type: REVIEW ARTICLE
Citation: Yamamoto T (2020) Opinion: The Collaboration of Adrenal Cortex and Medulla from Clinical Viewpoint. Int J Endocrinol Metab Disord 6(1): dx.doi.org/10.16966/2380-548X.166
Copyright: © 2020 Yamamoto T. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
Received date: 25 Mar, 2020
Accepted date: 20 Apr, 2020
Published date: 27 Apr, 2020
