Abstract
Background: Acne vulgaris is a chronic inflammatory disease that affects the majority of the population at some point in their lifetime. Acne pathogenesis is multifactorial with four primary contributors that play a pivotal role in the formation of acne lesions: inflammation, androgeninduced sebum production, abnormal keratinization, and bacterial colonization. Recent studies have demonstrated the anti-inflammatory, anticarcinogenic, anti-diabetic, and anti-lipid properties of certain Polymethoxylated Flavones (PMF) derivatives.
Objective: To study the anti-inflammatory activity and inhibition of lipid production by nobiletin and a mixture of PMF in three-dimensional (3D) epidermal models and sebocytes.
Methods: Oil Red O staining of primary human sebocytes treated with nobiletin and Mixture PMF was used to test the Inhibition of lipid production. Microarray analysesin three-dimensional (3D) epidermal tissue models were used. QuantiSig® luciferase reporter assay was used to study the effect of nobiletin and Mixture PMF on activation of nuclear receptor Peroxisome Proliferator-Activated Receptor (PPAR) (α, β/δ, γ).
Results: Nobiletin and Mixture PMF significantly reduced sebum production in primary human sebocytes and inhibited PPAR activation. The known anti-acne compound retinoic acid (RA) stimulated PPAR activation, suggesting possible different sebum reduction pathways. Nobiletin and Mixture PMF down-regulated PPAR gene expression and inhibited cytokines and immune receptors involved in the inflammatory response. Nobiletin downregulated cytochromes P450 cluster of CYP family of genes: CYP4F22, CYP4B1, CYP4Z2P suggests inhibition of lipid production by nobiletin via reduction of hormonal stimulation.
Conclusions: Nobiletin and Mixture PMF inhibited the inflammatory cascade, androgenic pathway in 3D human epidermal tissue models compared to the vehicle-treated cells. Nobiletin treatment of epidermal tissues reduces lipid accumulation in primary human sebocytes, inhibits PPARδ activation compared to the vehicle-treated cells.
Keywords
Polymethoxylated flavones; Microarray analyses; Cytochromes P450
Abbreviations
PPAR: Peroxisome Proliferator-Activated Receptor (α,β/δ,γ); Mixture PMF: Polymethoxylated Flavones Mixture; NF-κB: NFkappaB; MMP: Matrix Metalloproteinase; RA: Retinoic Acid
Introduction
Acne is the most common skin condition in the United States of America, affecting 80% of people between 11 and 30 years of age [1,2]. It equally affects genders, skin types. By age 21, 90% of men and 80% of women will have acne vulgaris [3]. Acne can persist in affected patients throughout adulthood, or appear de novo at any time and does not substantially decrease until after age 44 (P<0.001) [1]. The acne treatment field is witnessing a shift from formulated drugs limited to reducing sebum production to compounds targeting specific pathways of the immune system, which could be administered safely and available over the counter.
Acne vulgaris is a chronic inflammatory disease: factors in these conditions are the increased sebum production, hypercornification, and abnormal differentiation of keratinocytes of the upper part of the follicle. Hormones are other contributing factors that influence the severity of acne. Additionally, skin resident bacteria play a role in disease pathogenesis. The main three families of bacteria on the skin surface belong to Corynebacteria, Propionibacteria, and Staphylococci [4]. The interplay between members of this cutaneous microbiota and innate immunity is essential for the maintenance of healthy skin. Thus acne pathogenesis is multi factorial with four primary factors that play a pivotal role in the formation of acne lesions: inflammation, androgen-induced sebum production, abnormal keratinization, and bacterial colonization of Cutibacterium acnes in the pilosebaceous unit [5].
Recent evidence suggests inflammation as the inciting factor rather than activation of sebaceous glands, with proof of significant inflammatory factors surrounding the pilosebaceous unit even in clinically normal skin units in acne patients [6]. A new understanding of how Cutibacterium acnes (C. acnes) induce the inflammatory cascade has provided a new paradigm for the management of acne. Recognition of microbes, such as C. acnes, by the innate immune system is the body’s first line of defense against Pathogen-Associated Molecular Patterns (PAMPs) and Damage-Associated Molecular Patterns (DAMPs). C. acnes triggers innate immune response via the activation of toll-like receptors 2 and 6 (TLR2/6) and importantly the inflammasome [7]. The inflammasome, a caspase 1-activating cytoplasmic complex induces the secretion of crucial proinflammatory cytokines, including tumor necrosis factor α (TNF-α), IL-1, IL-6, IL-8, and IL-12 [8].
It is reported that nobiletin modulates a wide array of signalling pathways involved in the immune response, supporting its potential for use against inflammatory diseases such as acne. In contrast to RA, the main anti-acne treatment, PMFs derived from citrus peels prevented UVB-induced photo-inflammation by suppressing the expression of COX-2 in human keratinocytes [3,9]. Recent studies have demonstrated the anti-inflammatory, anti-carcinogenic, antidiabetic, and anti-lipid properties of certain PMF derivatives [10-13].
Our data show that nobiletin treatment of epidermal tissues or sebocytes inhibits a) the inflammatory cascade; b) androgenic and estrogenic pathways; c) lipid production and d) PPARδ activation.
Materials and Methods
Reagents
Nobiletin (N; Deep Sense LLC, USA) stock solution was prepared in DMSO and tested at 0.02mg/mL final concentration. 5-Demethylnobiletin (5-D-Nob; Deep Sense LLC, USA) stock solution was prepared in DMSO and tested at 0.01mg/mL and 0.005mg/mL final concentrations. Rosiglitazone (Rgz; Cayman Chemical, MI, USA) stock solution was prepared in DMSO and was used at concentration 9μg/ml as a positive control for PPAR activation. Retinoic acid (RA; Sigma, MO, USA) stock solution was prepared in DMSO and tested at 1μM final concentration in cell culture medium.
Cell culture
Primary human epidermal keratinocytes derived from the foreskin of a single donor were purchased from PromoCell (Heidelberg, Germany) and grown in supplemented Epilife medium (Thermo Fisher Scientific, NJ, USA). Differentiated 3D epidermal tissues were cultured on tissue inserts (Thermo Fisher Scientific/Nunc, CA, USA) at SkinAxis, LLC (Cedar Knolls, NJ, USA). The cells were submerged in supplemented Epilife growth medium for 48 hours and then grown in keratinocyte defined medium (KDM, SkinAxis, NJ, USA) at the air-liquid interface for 8 days. Keratinocytes and cultured epidermal tissues were maintained at 37°C with 5% CO2 and ~95% humidity.
3D epidermal tissues were topically treated with 25μl of compounds, positive or negative controls for 24 hours. After incubation, tissues were washed in PBS (Thermo Fisher Scientific), and total RNA was extracted using a Qiagen RNeasy kit (Qiagen, CA, USA) according to manufacturer’s instructions.
Cytotoxicity assay
Cytotoxicity was evaluated using the Cell Titer 96 Aqueous One (Promega, WI, USA) basic test according to the manufacturer’s instructions. Keratinocytes were seeded in a 96-well plate, cultured overnight, and then incubated for 24 hours in the presence of the compounds. Untreated and 5% DMSO-treated cells were used as a negative and positive control, respectively. All treatments were performed in triplicate. Treatments with compounds resulting in the inhibition of viability of more than 20% of the negative control values were considered cytotoxic.
Testing expression of PPAR promoter with luciferase reporter in response to nobiletin treatment
Cultured primary human epidermal keratinocytes were infected with VSV-G envelope pseudotyped lentivector carrying the luciferase reporter gene under the control of the PPAR , β/δ, γ) promoter (SkinAxis QuantiSig® system; SkinAxis, NJ, USA). Transduced keratinocytes were maintained at 37°C with 5% CO2 and 95% humidity for 24 hours. Treatment with different concentrations of compounds was done for 24 hours followed by luciferase and protein concentration assays.
Treatments of cultured epidermal tissues
Epidermal tissues (four biological replicates for each condition) cultured in tissue inserts (Thermo Fisher Scientific/Nunc) were topically treated with 25µl of compounds, positive or negative controls and addition of the treatments to cell culture medium. After 24-hour incubation, tissues were washed in PBS (Thermo Fisher Scientific), and total RNA was extracted using a Qiagen RNeasy kit (Qiagen, CA, USA) according to the manufacturer’s instructions. RNA concentration and purity were determined using a Nano drop IMPLEN spectrophotometer.
RNA quality control and microarray analysis
The integrity of total RNA samples and RNA quality was confirmed by Advanced BioMedical Laboratories (Cinnaminson, NJ, USA). A proprietary algorithm that takes several QC parameters into account (e.g. 28S/18S peak area ratios, unexpected peaks in the 5S region, etc.) was used to calculate the RNA Integrity Numbers (RIN). A RIN number of 10 indicated perfect RNA quality; a RIN number of 1 indicates degraded RNA. RNA with a RIN number ≥ 8 is of sufficient quality for gene expression profiling experiments. The RIN number for all RNA samples was >8. Microarray analysis was performed using Affymetrix Human Clariom D array processing by Advanced BioMedical Laboratories (Cinnaminson).
Statistical analyses
The differential gene expression was obtained using a threshold of 0.05 for statistical significance (p-value) and a log fold change of expression with an absolute value of at least 0.6. The microarray data herein presented reached statistically significant nominal p values. Gene expression was further analyzed using Differential Expression Analysis and Pathway Analysis (iPathway Guide; [14] by Advaita Bioinformatics Services (Plymouth, MI) in the context of pathways obtained from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (Release 84.0+/10-26, Oct 17) [15], gene ontologies from the Gene Ontology Consortium database (2017-Nov 6) [16], miRNAs from the miRBase (Release 21) and TARGETSCAN (Targets can version: Mouse:7.1, Human:7.1) databases [17,18] and diseases from the KEGG database (Release 84.0+/10-26, Oct 17) [19].
Data are presented as mean ± SD. Two-tailed t-test or two-way ANOVA with Bonferroni post-test were used for statistical analyses using Graph Pad Prism software. P value<0.05 was considered statistically significant.
Results
Evaluation of intracellular lipids in primary human sebocytes treated with nobiletin or Mixture PMF
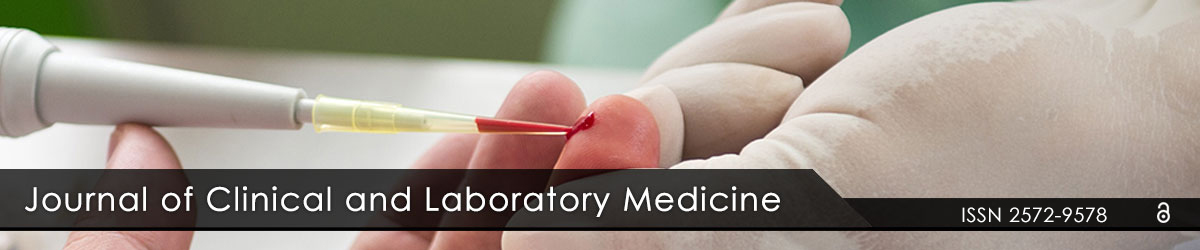
Preliminary experiments were carried out to determine the noncytotoxic range of nobiletin and a mixture of Nobiletin and Tangeritin (Mixture PMF) which has solubility and permeation similar to Nobiletin (data not shown). Normal human sebocytes (ZenBio, Research Triangle Park, NC) were cultured in the presence of noncytotoxic concentration (10μg/ml) of nobiletin or Mixture PMF for 72 hours before their effect was assessed by measuring the intracellular lipids content following Oil Red O staining (Figure 1A). Oil Red O staining was quantified by using Image J for the percentage of area stained positively for Oil Red O. Quantitative analysis revealed that the percentage of area stained positively for Oil Red O was significantly reduced in treated cells compared to cells cultured in growth medium (Figure 1B).
Figure 1A, 1B: Reduction of Oil Red O staining in primary human sebocytes treated with nobiletin and mixture of PMF. 1A) Microscopy detection of lipid droplets in sebocytes treated for 72 hours with either with medium control, 1µM retinoic acid (RA) positive control, 25µM Nobiletin, or 10µg/ ml Mixture PMF (Mix). Arrows indicate lipid droplets in cells. Original magnification: 200X. 1B) Quantitative analysis using Image revealed that the percentage of area stained positively for Oil Red was significantly lower in treated cells than in growth medium cultured cells. One-way ANOVA: The data are expressed as mean ± SD, n=6, *P<0.05 vs. growth medium treated cells.
Inhibition of PPARs by nobiletin and Mixture PMF
Nobiletin and Mixture PMF cytotoxicity curves were first determined in primary human epidermal keratinocytes derived from the foreskin of a single donor (PromoCell, Heidelberg, Germany) and grown in EpiLife supplemented medium (Thermo Fisher, NJ, USA). The nuclear receptor peroxisome proliferator-activated receptor gamma (PPARγ) controls the expression of different genes involved in the regulation of lipid pathways.
Studies in human sebocytes have raised the possibility that PPARγ modulation can lead to alteration of lipid production and may be useful in the treatment of acne [20]. The effect of nobiletin and Mixture PMF on the PPAR pathway was assessed using SkinAxis’ QuantiSig®
reporter system in which primary human keratinocytes are engineered with a luciferase gene under the control of the PPAR (α, β/δ, γ) promoter.
Nobiletin showed a decrease of reporter expression as compared to control vehicle-treated keratinocytes by 58% and 63% in the presence of PPAR activator -roziglitazone respectively indicating significant suppression of the PPARs activation (Figure 2). The mixture of PMFs at a final concentration of 10µg/ml lowered the reporter expression by 49% and 51% in the presence of rosiglitazone compared to control treated cells (Figure 2). As expected, significant stimulation of the reporter was observed with the positive control Rosiglitazone (151% Figure 2). RA at 1µM produced a significant stimulation (214%) of the reporter expression compared to the vehicle control (Figure 2). Our data on RA stimulation of the PPAR pathway are in agreement with other studies showing activation of PPAR by RA [21,22].

Figure 2: Nobiletin and Mixture PMF reduce PPARs activation. Activation of the PPAR promoter in cultured human keratinocytes treated as indicated for 24 hours. (The PPAR pathway activator Rosiglitazone (RGZ) at 25μM was used as positive control. The known anti-acne compound, Retinoic acid (RA) at concentration 1μM, was used as a reference, nobiletin (at concentration 25μM) and mixture Polymethoxyflavones (PMF) at 10ug/ml. All readings were normalized to total protein concentration and data are reported relatively to the expression of the reporter in the vehicle control (Vehicle negative control: 100%). One way ANOVA: *P<0.05; **P<0.01.
Microarray analyses of 3D human epidermal tissues treated with nobiletin
To evaluate the effect of Nobiletin (25μM), Mixture PMF (10μg/ ml), and RA (1μM) on gene expression profile in cultured human epidermal tissues (Figure 3) microarray analyses was used. Nobiletin and RA treatment affected the PPAR pathway by triggering different gene expression profiles. Microarray results confirmed our QuantiSig® data showing RA stimulation but Nobiletin and Mixture PMF downregulation of genes involved in the PPAR pathway. Moreover, the PPARδ gene was significantly down-regulated by Nobiletin and Mixture PMF treatment (-0.731 and -1.368 Log Fc) respectively), but was not affected by RA treatment. These data suggest that Nobiletin may trigger different sebum reduction pathways than RA.
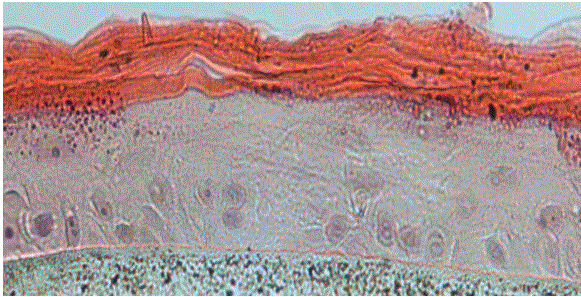
Figure 3: Hematoxylin and eosin staining of three-dimensional epidermal tissue.
Sato et al. 2007 showed that a mechanism of suppression of sebum production by Nobiletin includes inhibition of Diacylglycerol Acyltransferase (DGAT) [23]. Our microarray data confirmed the reduction of the DGAT gene expression not only by Nobiletin (-1.241) and Mixture PMF (-1.417), but also by RA (-1.1), suggesting that the inhibition of DGAT gene expression is common to Nobiletin, Mixture PMF as well as RA. Importantly, we also showed that Nobiletin downregulated the expression of cytokines and immune receptors involved in inflammation (Table 1).
| Gene Symbol |
Log Fc |
p-value |
| IL36RN |
-2.567 |
1.000e-6 |
| IL7R |
-1.618 |
2.197e-6 |
| IL1RN |
-1.618 |
1.056e-4 |
| IL1R2 |
-1.604 |
5.065e-4 |
| IL22RA1 |
-0.0.796 |
7.745e-4 |
| IL20RA |
-0.795 |
0.002 |
| IL37 |
-0.882 |
0.004 |
| IL36G |
-0.664 |
0.004 |
| PPIL6 |
0.795 |
0.005 |
| NF-κB |
-0.629 |
0.006 |
Table 1: Treatment of epidermal tissues with nobiletin down regulated
cytokines, receptors and nuclear factor involved in inflammatory
response.
Cytochrome P450 (CYPs) regulate skin homeostasis and metabolic barrier of the skin through activation or inactivation of different pathways. Cytochrome P450 proteins catalyze many reactions involved in drug metabolism, including the synthesis of cholesterol, steroids, and other lipids [24].
The local production of androgens and estrogens from steroid precursors is initiated by locally expressed CYPs [25].
Our microarray studies revealed that Nobiletin and Mixture PMF down-regulated a cluster of CYP genes in the cytochrome P450 family: CYP1A1, CYP4F22, CYP2S1, CYP1B1, CYP34, CYP39A1 (Table 2) suggesting that Nobuletin could decrease lipid synthesis. Local production of androgens and estrogens in the skin is regulated by a combination of CYPs (CYP11A1, CYP11B1, CYP17A1) and steroid dehydrogenases. Down-regulation of indicated CYPs by Nobiletin treatment was not statistically significant. However, nobiletin treatment of epidermal tissues down-regulated the estrogen signaling pathway by reducing the expression of genes involved in this pathway (Table 3).
| Gene Symbol |
Log Fc |
p-value |
| CYP1A1 |
-1.679 |
1.619e-6 |
| Cyp4F22 |
-2.331 |
1.000e-5 |
| CYP2S1 |
-0.718 |
8.368e-4 |
| CYP1B1 |
-1.618 |
1.056e-4 |
| CYP3A4 |
-0.655 |
0.011 |
| CYP39A1 |
-0706 |
0.001 |
Table 2: Nobiletin treatment of epidermal tissue down-regulated
Cytochrome P450 subfamily gene members.
| Gene Symbol |
Log Fc |
p-value |
| KRT10 |
-2.743 |
3.837e-6 |
| KRT23 |
-1.822 |
3.57e-6 |
| MMP9 |
-1.113 |
4.618e-4 |
| HBEGF |
0.783 |
1.359e-4 |
| MAPK3 |
-1.028 |
1.548e-4 |
| MMP2 |
-0.922 |
8.509e-4 |
Table 3: Nobiletin treatment of epidermal tissue down-regulated genes
involved in estrogen signalling pathway.
Discussion
Human skin is the largest body organ and is considered as the first line of defense and barrier against infections caused through the skin. Acne vulgaris is a very common inflammatory disease occurring primarily on the face and also rare on the upper arms, trunk, and back, which is caused by different bacterial species [4]. Despite advancement in science, the development of natural herbal treatments for skin disorders becomes an essential contributor for not only managing the side effects of conventional medicines but also more effectively treating skin disease. Herbal extracts and their phyto medicines make a fundamental contribution to human health care.
PMFs are reported to have several biological effects in vivo and in vitro, including antioxidant, anti-inflammatory, and anti-tumorigenic activities [13,26,27]. Our microarray data are in agreement with published reports on the anti-inflammatory activity of nobiletin [28]. We showed that the Nobiletin treatment of epidermal tissues downregulated the expression of cytokines and immune receptors involved in inflammation. NF-κB activation plays a key role in innate immunity and inflammation, and inhibition of NF-κB has been considered as a highly promising anti-inflammatory therapy. We found that the nobiletin and mixture PMFs strongly down-regulated NF-κB and IL-36RN gene in addition to other key cytokine genes resulting in lowered IL-RA proteins responsible for causing inflammatory skin diseases such as acne and psoriasis.
In this study, we examined in vitro effects of nobiletin on oil production compared to RAs. In-vitro studies with primary human sebocytes were used to determine the effect of nobiletin and PMF mixture on oil production. Sebocytes, despite being of epithelial origin and expressing the number of hormone receptors, are involved in lipid synthesis and metabolism, a function that is normally found in adipocytes. The sebaceous gland can be considered both as a target for hormones as well as an endocrine organ.
Our results are in agreement with the published report from Sato et al., on inhibition of lipogenesis by nobiletin. One of the mechanisms of suppression of sebum production by Nobiletin reported by Sato et al., was considered inhibition of Diacylglycerol Acyltransferase (DGAT) [23] However, while our microarray data confirmed the reduction of the DGAT gene expression not only by Nobiletin and Mixture PMF but also by RA, suggesting that the inhibition of DGAT gene expression is common to Nobiletin, Mixture PMF as well as RA [23].
We have shown that the Mixture PMF and nobiletin inhibition of lipid production in primary human sebocytes is different from the pathways triggered by RA. We discovered that, unlike RA, the PMFs inhibit the PPARδ signaling pathway, while RA-stimulated PPAR expression and has no effect on PPARδ. The graphical summary of the results is presented in figure 4.
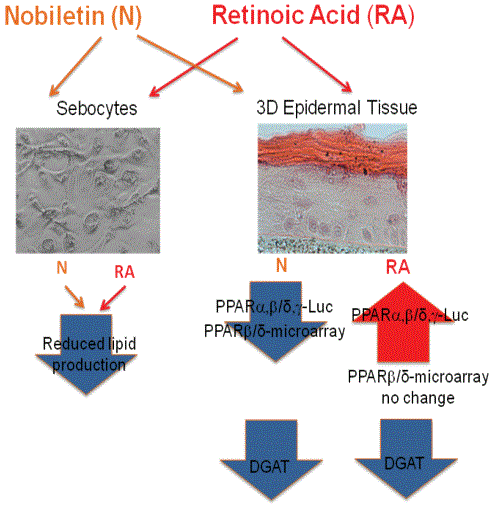
Figure 4: Graphical summary of the work.
PPARδ regulates a multitude of physiological processes associated with lipid metabolism, inflammation, immune response, and proliferation. The PMFs therefore can help modulate antiinflammatory properties that target keratinocytes in the epidermis and sebocytes in the superficial dermis to effectively treat acne vulgaris. They also open opportunities for treatment of other inflammationmediated skin disorders including psoriasis and melanoma.
The metabolic barrier of the skin is composed of cytochromes P450 (CYPs) that regulate its homeostasis through activation or inactivation of different pathways. The cytochrome P450 proteins are monooxygenases that catalyze many reactions involved in drug metabolism, synthesis of cholesterol, steroids, and other lipids [24]. The local skin steroidogenic system is composed of locally expressed CYPs involved in the local production of androgens and estrogens from steroid precursors [25]. Our microarray studies revealed that Nobiletin and Mixture PMF down-regulated not only a cluster of CYP genes in the cytochromes P450 family but also genes involved in the estrogen pathway, suggesting that Nobuletin could inhibit local production of androgens and estrogens and thus decrease lipid production possibly via the local inhibition of the hormonal pathway.
Data Availability Statement
All the data used to support the findings of this study are included within the article.
Funding Information
This work was supported by the funding from SkinAxis LLC.
Competing Interests
The author confirms that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
Ethics Approval and Consent to Participate
Studies reported in this manuscript do not involve animals or human subjects; human cell lines were obtained from indicated companies with de-identified donor information.
Acknowledgments
We are grateful to Dr. Jessica Langer for advice and critical reading of the manuscript.