Introduction
Adolescence is a challenging time of voice change, normally, and in pathology, where lack of capability of voice production hinders social function. The understanding of normal hormonal changes is essential to help diagnose voice problems in pathology. Therefore, we tried to make an overview of hormonal- and adolescence research, focusing on where voice production science could be connected.
Technical methods for measuring voice production include high speed films combined with analysis programs, e.g. Glottal Analysis Tools (GAT). Convolution networks analysis of high speed films with 4000 pictures per second is used in the clinical setting as well as optical coherence tomography [1-3]. These methods with high amounts of input information should be related to hormonal and other biological measurements in adolescence, to help treat many developmental disorders that include voice production.
The aim of the overview is to confirm the state of the art, of the connections between exact measurement of development of voice production, and adolescence related hormonal development-to open for supplementary understanding of voice and brain function in pathology. Voice production measurements should be an integrated part in pathology e.g. genetic malformations including cochlear implants [4,5] as well supplement the arbitrary definition of pubertal voice break, as it is made in research even now a days by self-evaluation of clients.
Methods
With Medline, Embase and hand search the amount was small of hormonal research papers that included voice production studies in adolescence, as searched by the librarians at the Royal society of medicine, UK. Table 1 shows the search methods for the systematic review of “Voice, Hormonal and Gender Development”. The search resulted in 9 papers, therefore, we included our own intensive search of other papers on adolescence, often based on reference lists in studies of other subjects, that could be relevant for understanding development in adolescence even if the papers were without voice measurements.
| Set# |
Searched for |
Results |
| S8 |
s7 not ti (transgender* or “trans men”) |
214* |
| S7 |
(s1 or s2 or s3) and (s4 or s5 or s6) and (la (english) or abany (yes)) and pd (2010-2020) |
222 |
|
S6 |
MJEMB.EXACT (“sex hormone”) OR MJEMB.EXACT (“hormone”) OR MJEMB.EXACT.EXPLODE (“hormone action”) OR MJEMB.EXACT.EXPLODE (“sex hormone”) |
327531 |
| S5 |
MJMESH.EXACT.EXPLODE (“Hormones”) OR MJMESH.EXACT.EXPLODE ("Sexual Development”) |
910938 |
|
S4 |
ti,ab((hormone [*1] or hormonal or testosteron* or [*2] strogen* or progesteron*) near/5 (develop* or puberty or chang* or increase* or decreas* or maturation or adolescen*)) |
264432 |
| S3 |
MJEMB.EXACT (“vocal cord”) OR MJEMB.EXACT (“voice”) |
18716 |
| S2 |
MJMESH.EXACT.EXPLODE (“Voice”) OR MJMESH.EXACT (“Vocal Cords”) |
14166 |
| S1 |
ti,ab ((voice[*1] or vocal*) near/5 (chang* or develop*)) |
12589 |
Table 1: Search Strategy
Databases: Embase®, MEDLINE® (run 24/03/20)
*The search strategy retrieved a number of references that were then hand searched to find the most relevant. The details of 9 references have been provided in accordance with your original request.
Results
Studies of brain and genetics related to hormonesand voice production in adolescence
Of the 9 papers found by the systematic search for Voice, Hormonal, and Gender Development, there were only 4 usable papers [6-9]. Two other papers were used as adjacent studies [10,11].
Our subsequent hand search was mostly made, using reference lists from papers on adolescence. These papers on hormonal development did not include the development of voice production, but since the systematic search returned limited results (9 papers), these were used to illustrate the problem surrounding voice production.
The brain development of voice production related areas in adolescence is mostly genetic. It all begins from the nasal placode wherefrom GnRH (gonadotropin releasing hormone) neurons migrated to hypothalamus [12,13] as shown in Figure 1.
Figure 1 shows the central activation of GnRH neurons by leptin, considered necessary for normal pubertal development. The nasal placode, and development of GnRH neurons have been considered a causal mechanism as stimulating factors on KNDy neurons (Kisspeptin, neurokinin B, dynorphins). The genes involved are included in the figure. The neuropeptide alfa-MSH plays a key role in energy homeostasis by mediating the action of leptin and may have a central role to the metabolic control of puberty. From the hypothalamus, at the medio basal area, the arcuate nucleus (ARC), the pituitary gland is stimulated, also by the GnRH-PG (Gonadotropin Releasing Hormone-Prostaglandin), the reaction depending on the GnRH-R-(Receptors). Follicle stimulating and luteinizing hormones stimulate the ovaries in girls, and in boys the production of Leydigcells´ production of testosterone, stimulating puberty. In girls the Anti-Müller hormone (AMH) for egg reserve and inhibin level of hindering FSH production in the pituitary are regulated in balance with E2 (oestradiol), stimulating puberty. Extrinsic factors (EDC) and body mass index (BMI) have an impact on the development.
McCarthy [14] described in her book, the development of sex and the brain in animals in details but admits that animal studies are not directly useable in voice studies of humans. This is also the case for Perrodin C, et al. [15], Zaqout SI and Al-Hussain SM [16], Xie Y and Dorsky RI [17] discuss the development of brain incl. hypothalamus across vertebrate species. The knowledge of human subcortical development is summarized by Abbott, et al.[18]
Wierenga et al. [19] show that changes in testosterone level were related to development of pallidum, accumbens, hippocampus and amygdala volumes in males and caudate and hippocampal volumes in females. The modeling interaction between pubertal maturation and chronological age seemed to be sex specific. Etchell A, et al. [20] admit that sex differences in childhood language development is unclear-in a review of 46 published studies they conclude that sex differences may be more prominent during certain developmental stages due to different rates of maturation between the sexes. They admit that more research is needed to understand the influence of sex hormones and developmental stages. They notice that sex differences in brain structure and function do not necessarily lead to differences in task performance, and evidence for sex differences of development are limited, when present, sex differences often interact with a variety of factors such as age and task.
The important definition of puberty stages 1-5 is referring to Marshall and Tanner with voice change in boys between stage 2 and 4 [21]. Styne DM [22] made a thorough overview of puberty phenomena referring to Tanner and including genetics, hormones etc, voice is not discussed. This is also the case in the survey of puberty by Busch AS, et al. [6]. The author state here that puberty cannot be perceived as a solitary event, they discuss basic genetic changes, and hormonal changes as well as brain changes as earlier referred to, in this paper. The concepts are of 1. adrenarche (of production DHEAS and and rostenedione in the adrenals) 2. thelarche, breast development 3. menarche, beginning of menstruation, 4. pubarche, pubic hair development, 5. godanarche, secondary sex characteristics. Styne DM [22] also discusses prepubertal values of serum hormone binding globulin (SHBG) and oestron/oestradiol with general guidelines for prepubertal values. They point out that the pubertal stages are determined by hormones, of which most of the circulating oestradiol and testosterone is associated with SHBG and that prepubertal boys and girls have equal concentration of SHBG.
The mutational voice is between 12, 5 and 14 years of age. The vocal folds in males at 16 years of age are 18-24 mm long with a fundamental (F0) of 130 Hertz. In girls, 16 years of age, the vocal folds are 15-20mm long and the fundamental frequency (F0) is 220-225 [23]. At 6-12 years the vocal folds have two layers, at 16 years the vocal folds have three layers, which is documented with optical coherence tomography [24]. Table 2 shows the relations between voice parameters and androgens in boys and table 3 shows the relation between voice parameters and androgens/oestrogens in girls.
| Age |
(years) |
8.7-12.9 |
13.0-15.9 |
16.0-19.5 pr yr. |
% change |
| No of boys |
|
19 |
15 |
14 |
|
| Serum testosterone |
(n mol/l) |
0.54 |
10.5 |
18,9 |
68 |
| Dihydrotestosterone |
(n mol/l) |
0.18 |
1.21 |
1.57 |
37 |
| Free testosterone |
(n mol/l) |
0.007 |
0.14 |
0.33 |
77 |
| Sexual hormone binding globulin |
(n mol/l) |
134 |
66 |
45 |
-16 |
| Delta 4 androstene dione |
(n mol/l) |
0.54 |
1.17 |
2.5 |
24 |
| Dehydro epi andro sterone sulfate |
(n mol/l) |
1400 |
4100 |
5900 |
25 |
| Testis volume |
(ml) |
2,3 |
13 |
20 |
36 |
| Fundamental frequency |
(Hz) |
237 |
184 |
125 |
-11 |
| Voice range |
(semitones) |
3.7 |
4.8 |
5 |
3.9 |
| Phonetogram area |
(cm2) |
19 |
28 |
34 |
9.2 |
| Lowest biological tone |
(Hz) |
158 |
104 |
72 |
-12 |
Table 2: Shows the relations between voice parameters and androgens in boys.
Geometrical average of hormonal, pubertal and vocal parameters for boys grouped according to pre-pubertal, pubertal, post-pubertal age and the annual change in these parameters in %. (Phonetogram area: 1cm2 =32 semitones × dB(A)) [49].
| Age |
(years) |
8.7-12.9 |
13.0-15.9 |
16.0-19.8 |
Significance |
| Total number |
|
18 |
12 |
11 |
|
| Oesterone (E1) |
pmol |
57 |
104 |
123 |
** |
| Oestradiol (E2) |
pmol |
73 |
135 |
108 |
|
| Total testosterone |
nmol |
0.5 |
0.76 |
0.94 |
|
| Free testosterone |
nmol |
0.006 |
0.037 |
0.009 |
|
| Oesterone sulphate (E1SO4) |
pmol |
732 |
1924 |
2342 |
** |
| DHEAS |
nmol |
3210 |
3700 |
7200 |
** |
| Androstendione |
nmol |
1.44 |
3.28 |
3.43 |
* |
| Sex hormone binding globulin (SHBG) |
nmol |
153 |
130 |
123 |
|
| Menarche |
|
+4 |
+9 |
+11 |
|
| Pubic hair stage |
|
1-4 |
2-5 |
4-6 |
|
| Mamma development stage |
|
1-4 |
2-5 |
5 |
|
| Fundamental frequency in continuous speech |
Hz |
256 |
248 |
241 |
|
| Tone range in continuous speech |
Semitones |
3.7 |
4.2 |
5.2 |
** |
| Tone range in singing |
Semitones |
23 |
30 |
38 |
|
| Phonetographic area |
cm2 |
17.3 |
21.8 |
28.3 |
** |
| Phonetogram lowest tone |
Hz |
166 |
156 |
145 |
* |
| Phonetogram middle tone |
Hz |
429 |
409 |
413 |
|
| Phonetogram highest tone |
Hz |
1136 |
1105 |
1263 |
|
Table 3: Shows the relation between voice parameters and androgens/oestrogens in girls.
Geometrical averages of hormonal, pubertal and vocal parameters for girls grouped according to pre-pubertal, pubertal, post-pubertal age. The relative standard deviation lay between 11% and 140%. (Significance of the differences between the groups: p < 0.01 xx; p < 0.05 x). Cm2 conversion factor: 1cm2=32 semitones × dB(a) [49].
Busch AS, et al. [6] still recently described “voice break” recurring at 13,6 years (13,5-13,8 years of age) The voice breaks was self-evaluated and corresponded to testis size 11,8 mm (4-20 mm) and genital stage 3 (stage 2-5). In pathology voice production related to puberty should be much more focused upon, as earlier mentioned e.g. in cochlear implants and in many other developmental disorders e.g. Turner syndrome [25,26].In adults there seems now to be an acceptance of the functional connectivity of among others, periaqueductal grey (PAG) with core limbic system and laryngeal cortico-motor structures during human phonation, divided in volitional and non-volitional phonation [27].
Figure 2 describes how input and ongoing voice modulation is from the posterior superior temporal gyrus (PSTG) as well as from the supplementary motor area (SMA) and Insula. Output from the primitive integrative vocalization centre (VOC) as shown by Penfield and Roberts [31] is described. Black lines in figure 1 are for the direct pathway via the corticobulbar pathway and the cerebellum (CBL) bilaterally. Grey lines show the pathway from VOC to the cingulate, Amygdala Nucleus (AM) Periaqueductal Grey (PAG) the pons and the reticular area in the medulla with input to Nucleus Ambiguous (NA) on both side of the brain. Human emotionally based vocalisation and volitional voice production have shown more integration than previously proposed- It should be noted that reflexive as well as learned voice production-not involuntary speech-have a common system.
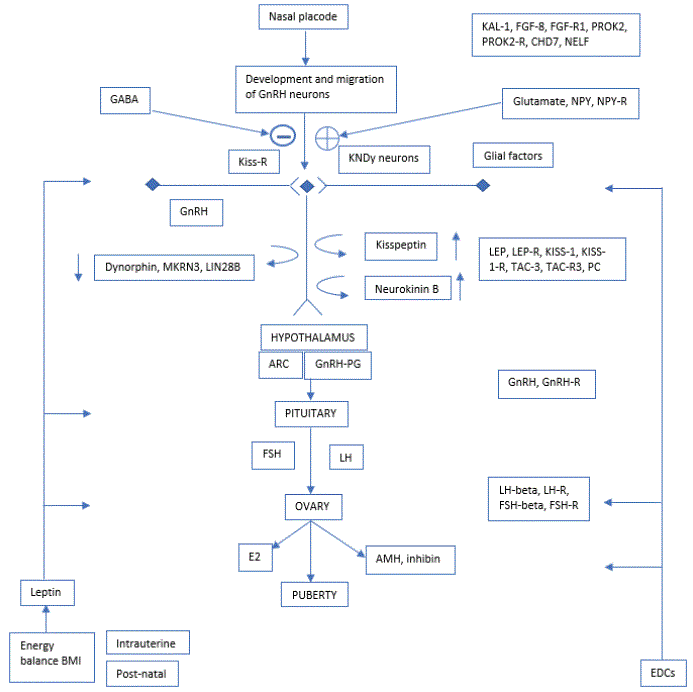
Figure 1: Presents an overview of the genes involved in puberty regulation with hypothalamus in the centre. The development starts from the nasal placode in the foetus with development and integration of GnRH neurons (gonadotropin releasing hormone expressing neurons).
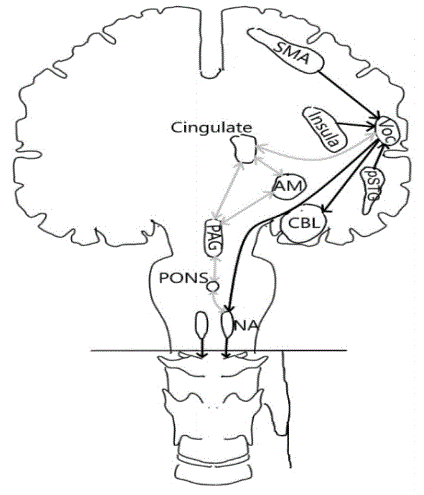
Figure 2: Input and ongoing modulation is from the posterior superior temporal gyrus (pSTG under the VOC) as well as from the supplementary motor area (SMA) and Insula. Output from the primary integrative vocalisation centre (VOC). Black lines are the direct pathway via the corticobulbar pathway and the cerebellum (CBL) bilaterally. Grey lines show the pathway from VOC to the cingulate, nucleus amygdalae (AM), periaqueductal Gray (PAG), the pons and the reticular area in the medulla with input to nucleus ambiguous (NA) on both sides of the brain. From there to the vagus nerve.
Holstege G, et al. [28] underline that only humans can speak because, via the lateral components of the volitional or somatic motor system, they are able to modulate vocalization into words and sentences by activation of the prefrontal area, PAG, and caudal medullary Nucleus Retro Ambiguous (NRA). NRA is the only cell group that has direct access to the motor neurons involved in vocalizations. On a planned large-scale chart for developmental changes in the brain, a focus will be on the pubertal period from 9-17 years of age, and structural plus functional activity will be compared with pubertal hormones and pubertal development [29]. Vasung L, et al. [30] conclude that it remains unclear how among others metabolic demands influence the development.
Acoustics related to hormones and voice productionin adolescence
There was a change of understanding of the human voice with the vowel research made by Peterson and Barney [31]. Studies have been made of the role of formant overtone maxima in children [32]. An important approach is the study of the development of resonances in speech and singing as well [33-35]. The sex hormone receptors in vocal folds have been focused upon by Nacci A, et al. [10] but were seldom found. The authors speculate that the changes of voice according to gender throughout life might be linked with a different expression of some growth factor in the laryngeal tissue and that this expression might in turn be influenced by hormonal variation. Interesting is a study from Sato K, et al. [36] showing that the vocal fold mucosa, unphonetic, without voice, two cerebral palsy children (7 and 12 years old) did not have a vocal ligament, lamina propria appeared as a uniform structure, vocal fold stellate cells synthesized fewer extracellular matrix substances such as fibrous protein and glycosaminoglycan.
Hormones and adrenarche
Adrenarche encloses the change of the outer layers of adrenals related to pubic and axillary hair. Guran T, et al. [37] measured the age and DHEA-S secretion in healthy children at adrenarche, where DHEA-S concentration over 108,4 nm/L (40 µg/mL) were regarded as adrenarche at 8 years of age in girls and 7 years for boys. Dörr HG, et al. [38] confirm that DHEA-S were higher in Turner girls without growth hormone therapy, but that the time of adrenarche were the same with or without hormone therapy. Whittle S, et al. [39] focus on adrenarche timing on brain function,in the study of early adrenarche defined by high DHEA-S independent of age of effects on brain function. 83 children, mean age 9,53 (SD 0,34) hereof, 43 female were examined, and a higher DHEA level was associated with decreased affect related activity in the mid-cingulate cortex in the whole sample, and in a number of cortical and sub cortical regions in females. Higher DHEA levels were also associated with increased externalizing psychological symptoms in females that were partly mediated by posterior-insula activation. The author suggests that timing of adrenarche is an important moderator of affect related brain function. Barendse MEA, et al. [40] confirm that adrenarche changes in the brain structures are prior to godanarche. In a longitudinal study of DHEA/ DHEA-S and testosterone on fMRI at 9 years, high DHEA suggested negative association with white matter microstructure. Higher values were also related to anxiety symptoms through an effect on amygdala and inferior frontal gyrus. Testosterone was related to the development of white matter.
Development of voice production should be much more exact than self-reported voice break to compare to other parts of body development and pathology [6]. There are many developmental disorders, where voice production is part of the handicap [5]. New focus on voice development was made in treated adrenal cortical tumours. In 9 adults and 10 adolescents female, F0 was reduced to 132 Hertz in 1 and F0 to 165/168 Hz respectively in two subjects. The majority had normal F0 , from 189 to 245 Hz [41].
Adjacent studies
There are some aspects of early development that influence the voice related hormonal situation in adolescent youngsters: Especially the “mini-puberty” around birth and growth hormones. A problem is that a lot of research on hormones is on primates/ non-humans, which means that even if some results are common- when it comes to human voice, they can probably not be used. For the “mini-puberty” around perinatal life it is important to understand that peripheral hormones in blood serum are regulators in a transitory hormone surge that is comparable in its extent to puberty [42-44]. Borysiak A, et al. [11] noted that F0 properties were correlated to average serum levels of bio available oestradiol (E2) (mean E2/SHBG and testosterone/mean testosterone across the second month of life. They interpreted the results as indication of E2 influence on viscoelastic properties of vocal folds.
In rats [8,9] a study was made on the role of the vocal folds, especially the extracellular matrix (ECM) in vocal fold lamina propria. The hyaluronic acid decreased in ovariectomized rats. The collagen-1 was lowered, this was also the case of collagen-3 later in the observational period. Elastin was less dense in the same rats. They suggest that the vocal folds are an oestrogen sensitive target organ.
The study of Hodges-Siemon CR, et al. [45] and her group on testosterone and vocal parameters in 91 adolescent male indicated that males in better energetic conditions (BMI-for-age residuals from Tanner-specific growth curves) have higher testosterone and lowered voice even controlling for age. Testosterone given to females is a wellknown risqué of voice lowering. Wuntakal R, et al. [46] refer to some voice effects of LHRH agonists given for ovarian cancer. Zacharin M [47] has an overview of treatment of secondary hypogonadism of treated severe illness in childhood and adolescents since many children now survive chronic illnesses.
Prediction by hormones of voice production change in adolescence
Shirtcliff EA, et al. [48] has made an overview of hormonal change in puberty where it was shown that a picture-based interview combined with testosterone, DHEA and physical exam gave better predictive values. SHBG had a predictive significance of F0 change in boys of p<0.05, in girls the widening of the tone range of the fundamental frequency in semitones (F0range) with electroglottography during reading of a standard text and E1So4 measureshad predictive values of p<0.05 [49].
Gaidano G, et al. [50] found that mean values of SHBG binding capacity, both for dihydrotestosterone and testosterone were significantly higher in prepubertal subjects. The binding capacity of SHBG is a result of a pool of proteins which modifies pubertal evolution. Rosner W, et al. [51] discus SHBG as a cell regulator, they demonstrate an additional mode of action on steroid hormones, one that does not require that the steroid interacts with a receptor. Kim MR, et al. [52] measured SHBG which decreases markedly during early puberty. They found that 50 nm/L in stage Tanner 2 were significantly different to Tanner stage 1 and that free androgen index (FAI=testosterone/SHBG) could even better differentiate the onset of puberty. The decrease of SHBG coincidence with a significant increase in total body weight and body mass index.
Simo R, et al. [53] describe how sex hormone binding globulin (SHBG) is produced and secreted by the liver into the bloodstream where it binds sex steroids and regulates their bioavailability, by limiting their diffusion into their target tissue. Further research is needed to elucidate the molecular mechanism that could explain the sexual dimorphism regarding the plasma SHBG levels. Laurent MR, et al. [54] comment that the in-vivo physiological role of circulating SHGB remains unclear, transgene mice expressing a human SHBG transgene were used in their study. SHGB is an interesting factor in prediction of voice change in puberty in Tanner stage 2-4 due to its’ multifactorial tissue effects.
Discussion
Results of the systematic library search gave only 4 studies that included hormonal and voice production in adolescence
The voice break was self-evaluated in the hormonal studies. On the other hand, voice production development in puberty in Phoniatrics 1 [5] is described without hormones. With the social demands including voice production (voice performance) e.g. multi-handicapped patients are in a bad situation. We have the tools of high speed films and GAT, deep learning, and optical coherence tomography to analyze detailed sound and vocal fold phenomena in large scales. It is possible to compare the huge data amount with deep learning. This could give a more nuanced understanding of deviant voice behavior. And optical coherence tomography could at the same time give information of deviant tissue function.
Fundamental frequency (F0), tone range and the lowest tone
In adolescence it is easy to measure the F0 in a well-defined situation, e.g. measured during reading of a standard text or with standard counting and relate to testosterone/oestradiol. It is not only the mean F0 that is changing, also the frequency variation is expanding (the tone range) as it can be seen in the voice profile [5]. The lower tone becomes lower, which we know, is related to androgens and oestrogens (Table 2 and 3) [49]. This is used by singing teachers in choirs for prediction of pubertal change of voice production.
Prediction of the change of voice production in adolescence
The measurement of testosterone and oestradiol correlate to the change of F0 at the age of 13-15.9 years. Serum hormone binding globulin (SHBG) is significantly a predicting factor of F0 change in adolescence in boys. This allows us to speculate whether the change of voice could be related to a broader symptom complex [49-54]. Another way to get further understanding of the hormonal control of voice production might be to relate, as a standard not only to puberty stages 1-5 but also to the time of appearance of adrenarche, pubarche, godanarche, menarche and thelarche. This could specify relations of hormones and voice production usable also in pathology [24].
Conclusion
A systematic search was made on Voice, Hormonal, and Gender Development, which unfortunately only returned 9 papers, of which 4 were related to the subject. Additional hand search resulted in papers on hormonal changes and adolescence, but without voice production. Some papers were informative. The normal genetic regulation of hormones in adolescence is presented. Hormonal development and especially the Serum Hormone Binding Globulin (SHBG) has a predicting role for the development of voice production in adolescence which is not fully understood. In the future combined hormonal and updated online high speed measures of voice production can give a much better understanding of voice production development-eventually combined with optical coherence tomography and analyzed with neural networks. Especially in pathology, mostly genetic multi-handicaps´ syndrome patients could be better understood.
Author Contributions
Conceptualization: MP. Data curation: MP, AOJ. Formal analysis: MP, AOJ. Funding acquisition: MP. Methodology: MP, AOJ Visualization: MP, CL, AOJ. Writing – original draft: MP. Writing review & editing: MP, CL, AOJ.
