
Figure 1: Frequent use of selected tobacco products among U.S. middle and high school students who currently used each tobacco productNational Youth Tobacco Survey, 2017-2018 [48].
Allie Karges1 Mohammed S Inayat2 Vincent S Gallicchio3*
1Department of Psychology, College of Behavioral, Social and Health Sciences, Clemson University, Clemson, SC, USA*Corresponding author: Vincent S Gallicchio, Department of Biological Sciences, 122 Long Hall, College of Science, Clemson University, Clemson, SC USA 29627, USA, E-mail: vsgall@clemson.edu
Nicotine, primarily found in tobacco, exerts a profound impact on stem cells, disrupting their proliferation, differentiation, and migration when exposed at high doses or used chronically. Stem cells hold significant promise in the field of regenerative medicine and disease management, making it crucial to understand how nicotine affects their therapeutic potential. In tandem, a growing epidemic of nicotine addiction among adolescents poses a public health threat, as it not only jeopardizes their well-being but also undermines the efficacy of stem cell-based treatments. This paper reviews the effects of nicotine on various types of stem cells and discusses the alarming rise in adolescent nicotine use, emphasizing the importance of addressing this issue to preserve the future of stem cell therapies.
Nicotine; Stem cells; Pulmonary/respiratory dysfunction
AKT: Protein Kinase B; BM: Bone Marrow; CD31: Platelet Endothelial Cell Adhesion Molecule1; CD144: Integrin αII; COPD: Chronic Obstructive Pulmonary Disease; CSE: Cigarette Smoke Extract; EC: E-Cigarette; EMT: Epithelial Mesenchymal Transition; EPC: Endothelial Progenitor Cell; ESC: Embryonic Stem Cell; HGF: Hepatocyte Growth Factor; HLI: Hindlimb Ischemia; HSPC: Hematopoietic Stem and Progenitor Cell; IL6: Inflammatory Marker; IL8: Inflammatory Marker; IPSC: Induced Pluripotent Stem Cell; IV: Intravenously; JUUL: Specific e-cigarette devices that JUUL Labs manufacture; LCN2: Transporter protein lipocalin-2; LKS: Lin-c-kit+Sca-1+; LT-HSC: Long Term Repopulating Hematopoietic Stem Cell; MMP: Mitochondrial Membrane Potential; MSC: Mesenchymal Stem Cell; mtDNA: Mitochondrial DNA; nAChR: Nicotine Acetylcholine Receptors; NF-kB: Nuclear factor kappa B; NK: Natural killer; NSC: Neural Stem Cell; SLAM: Signaling Lymphocyte Activation Molecule (LinCD48-CD150+); STAT3: Signal Transducer and Activator of Transcription 3; VEGFR: Vascular Endothelial Growth Factor Receptor
Two primary pathways exist through which individuals can encounter cigarette smoke [1,2]. The first is the act of inhaling mainstream smoke, often referred to as “first-hand” smoke, which predominantly affects active smokers. This process involves the deposition of nicotine and hazardous smoke-related chemicals within the oral cavity or inhaling into the lungs. Subsequently, these substances are absorbed and transported through the bloodstream. “Second-hand” or side stream smoke refers to the latter pathway, which involves the smoke emanating from the lit end of a cigarette. This type of smoke exerts harmful effects on the lung function of the active smoker and any innocent bystander [2]. As a result, epidemiological studies have demonstrated a significant increase in the incidence of malignant and cardiovascular diseases in adults due to “second-hand” smoke exposure.
Research findings have indicated that nicotine, the primary addictive substance in tobacco, stimulates the growth, invasion, and development of blood vessels (angiogenesis) in cancerous cells while impeding their natural cell death (apoptosis) [3-5] examined the impact of nicotine exposure on angiogenesis and blood flow within the distraction regenerate of a rabbit model investigating mandibular lengthening. In the nicotine-exposed group, there was a significantly greater density of newly formed blood vessels compared to the control group. This is a problem because the formation of blood vessels supplies tumors with oxygen and essential nutrients. As a result, the tumor would enlarge, and cancerous cells may disseminate to other parts of the body. It is worth noting that previous research has indicated that nicotine can directly affect small blood vessels, inducing vasoconstriction and systemic vasoconstriction, ultimately leading to hypoxia [6,7].
A growing body of evidence suggests the both the innate and adaptive immune systems, primarily those of smokers, are largely affected by cigarette smoke [8-11]. Cigarette smoking has been linked to the induction of epithelial-mesenchymal transition (EMT) in breast and lung cancer, primarily through activating signaling pathways like STAT3, AKT, and NF-kB [12-14]. Activating these pathways can promote the formation of lung tumors by inciting inflammation due to these being crucial regulators of inflammatory responses [14].
Substantial evidence also indicates that nicotine significantly contributes to smoke-induced immunosuppression. This occurs by diminishing NK cell activity, suppressing T-cell proliferation, reducing macrophage antigen processing activity, and decreasing dendritic cell numbers [15-22]. Additionally, epidemiological studies have established a substantial association between cigarette smoking/ nicotine and an elevated risk of various cancers [23,24].
Nicotine is the primary ingredient in nicotine replacement therapies; gum and patches, which aid in quitting tobacco use [25]. While these therapies are considered safe and effective for cessation, the rise in e-cigarette popularity introduced another way of nicotine exposure [26]. Nicotine is considered a weak base with a pKa value of 8.0, and its absorption is more efficient in slightly basic environments, where it is less ionized. The oral cavity, where smoke exposure begins, is typically slightly acidic. Additionally, flue-cured cigarette smoke tends to be slightly acidic with a pH range of 5.5 to 6.0 [27]. These conditions in the oral cavity are not conducive to efficient nicotine absorption.
However, recent findings suggest that cigarette smoke may be more alkaline than initially believed, potentially improving nicotine absorption in the mouth [27]. In contrast, e-cigarette liquids tend to have a slightly more basic pH. As a result, nicotine delivered from these devices is thought to be absorbed more readily in the oral cavity [28]. Originally thought to be a safer alternative, recent clinical data suggest e-cigarettes might not help with quitting tobacco use and raise concerns about youth usage [29]. E-cigarette use among US high school students has surged from 1-2% in 2010-2011 to 27% in 2019 [30-32], possibly leading to increased tobacco use in adulthood. Although considered generally safe, nicotine and its byproducts can have adverse systemic effects, such as cardiovascular issues like atherosclerosis, ischemia, and myocardial dysfunction. Nicotine also affects vascular cells, promoting intimal thickening and atherosclerotic plaque progression [33].
Nicotine’s effects occur through nicotinic acetylcholine receptors (nAChR), consisting of five subunits [34]. Activating these receptors by acetylcholine or nicotine allows cation entry, leading to depolarization and downstream effects, including altered cellular behaviors like survival, proliferation, and apoptosis, due to calcium ion influx [35].
Although tobacco smoke is made of various toxins and chemicals, the addictive component stems from the presence of nicotine. Nicotine acutely activates brain regions like the prefrontal cortex, thalamus, and visual system, involving corticobasal ganglia-thalamic circuits [36]. This stimulation of central nAChRs leads to the release of several neurotransmitters, with dopamine being the primary one. Dopamine is released in areas such as the mesolimbic region, corpus striatum, and frontal cortex. The key to nicotine’s impact is the release of dopamine in the nucleus accumbens shell which is associated with drug-induced reward [37,38]. Nicotine also prompts the release of other neurotransmitters, including norepinephrine, acetylcholine, serotonin, GABA, glutamate, and endorphins, influencing various nicotine-related behaviors. The release of dopamine signifies a pleasurable sensation and plays a pivotal role in reinforcing the effects of nicotine [39]. Dopaminergic secretion reshapes brain circuitry related to learning, stress, and self-regulation, which all contributes to addiction and dependency [27,40,41]. Due to their heightened brain plasticity, adolescents and young adults are especially prone to nicotine receptor upregulation and addiction [42,43].
The surging popularity of electronic cigarettes has practically reversed five decades of declining nicotine usage among young individuals. Originally marketed as a smoking cessation aid for longtime adult smokers, vaping has become two to three times more prevalent among adolescents and young adults than older adults. This shift has been fueled by the emergence of vaping products, including a “mod pod” e-cigarette that utilizes its own cartridge-based nicotine produced by leading manufacturers like JUUL® which have dramatically altered the landscape of youth nicotine consumption. These products combine sophisticated technology-appealing designs and are heavily promoted through aggressive marketing and social media campaigns [44]. After the 2018 release of the Monitoring the Future report, the U.S. Surgeon General, Dr. Jerome Adams, and then FDA Commissioner, Dr. Scott Gottlieb, officially declared adolescent e-cigarette use an ‘epidemic’. Despite this nationwide attention, vaping among adolescents in the United States continued to increase. The swift proliferation of Juul pods, containing as much nicotine as 1.5- 2 packs of cigarettes, held approximately 70% of the U.S. e-cigarette market during that period [45]. The rise in use of these gadgets has corresponded with an outbreak of lung injuries linked to vaping, drawing national attention and causing considerable uncertainty about their respiratory risks [46].
Their increase in popularity can be seen through the surges in e-cigarette products bought by youth over past years. In a 2018 survey, 20.8% of high school students reported using e-cigarettes on more than one occasion in the preceding 30 days, marking a substantial increase from the 1.5% reported in 2011. Furthermore, 11.3% of high school students disclosed using two or more types of tobacco products, and among middle school students, the figure was 4.9%- nearly 1 in 20 [47]. In 2018, among high school students who were current users of various tobacco products, the prevalence of frequent use was as follows: 37.7% for smokeless tobacco, 27.7% for e-cigarettes, 23.1% for cigarettes, 15.8% for cigars, and 15.7% for hookahs (Figure 1). Notably, between 2017 and 2018, there was a significant 38.5% increase in frequent e-cigarette use among current e-cigarette users, rising from 20.0% to 27.7%. There were no significant changes in frequent use observed for other tobacco products.
Figure 1: Frequent use of selected tobacco products among U.S. middle and high school students who currently used each tobacco productNational Youth Tobacco Survey, 2017-2018 [48].
According to Gentzke, et al., middle school students in 2018 who were current users of various tobacco products, attributed frequent use rates as follows: 26.2% for hookahs, 22.7% for smokeless tobacco, 19.7% for cigarettes, 16.2% for e-cigarettes, and 15.0% for cigars. [48].
Exposure to nicotine during adolescence and young adulthood correlates with increased usage of other substances and has enduring impacts on the developing brain. Additionally, concerns arise from exposure to chemicals heavy metals, and the risks of acute injuries and poisonings associated with e-cigarette use. Despite contrary claims, unlike their potential benefits for older adults, there is no clear evidence that e-cigarettes serve as effective smoking reduction or cessation tools for young people, even among regular cigarette smokers trying to quit [44].
Unlike any other cells, stem cells represent a group of undifferentiated and are distinguished by their capacity for extensive proliferation. Typically, they originate from a single cell source and can differentiate into various cell types and tissues. These unique types of cells are derived from various sources, all exhibiting different degrees of potency.
Pluripotent cells are embryonic stem cells that originate from an embryo’s inner cell mass, while induced pluripotent cells result from adjusting somatic cells. These cells can differentiate into tissues belonging to all three germ layers: endoderm, mesoderm, and ectoderm. On the other hand, multipotent stem cells, like mesenchymal stem cells, can differentiate into tissues derived from a single germ layer, such as bone, cartilage, and adipose tissue [49].
The utility of stem cells extends to cellular therapy, replacing damaged cells or facilitating organ regeneration. Additionally, stem cells have significantly enhanced our comprehension of development and disease pathogenesis. Disease-specific cell lines can be cultured and utilized in drug development. Nonetheless, stem cell research faces challenges, including ethical dilemmas associated with embryonic stem cells, concerns about tumor formation, and issues related to rejection. Ongoing efforts are overcoming these limitations, holding the potential for substantial advancements in disease management [49].
Due to its multifarious effects, several types of stem cells (Figure 2) have been examined when determining the extent of nicotine’s threat on therapeutic potential, these include: circulating progenitor and stem cells, embryonic stem cells (ESCs), mesenchymal stem cells (MSCs), and induced pluripotent stem cells (iPSCs).
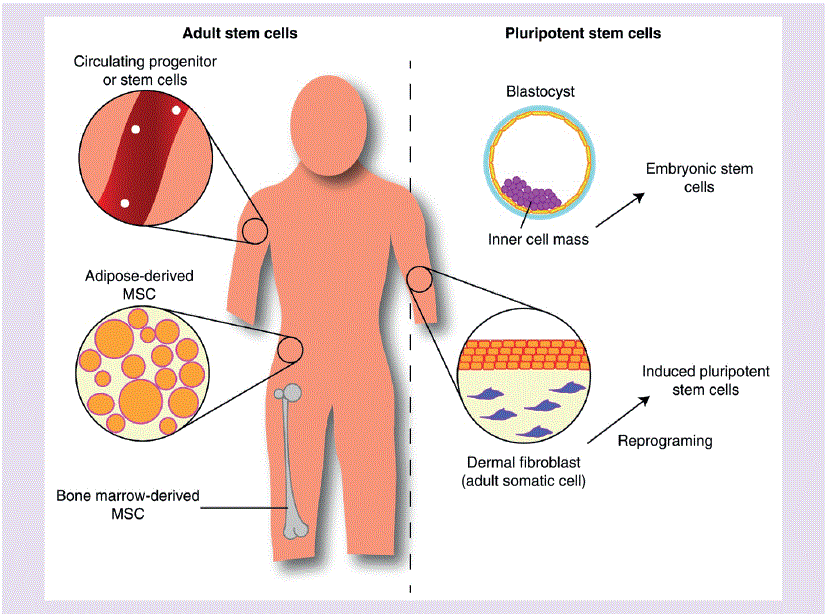
Figure 2: Illustration outlining various stem cell categories suitable for investigating nicotine exposure [33].
Endogenous circulating progenitor and stem cells originate in the bone marrow and circulate throughout the body, playing essential roles in various regenerative processes. In cardiovascular regenerative medicine, endothelial progenitor cells (EPCs) spike interest due to their capacity to develop into mature endothelial cells and facilitate angiogenesis in response to ischemic injury [50]. Given the rising incidence of ischemic conditions like coronary heart disease and peripheral artery diseases, EPCs hold significant promise in stem cell therapy.
EPCs have traditionally been identified based on the expression of cell surface markers like CD31, CD144, and VEGFR. However, a unique EPC marker is currently lacking because these markers overlap with those of endothelial cells [51]. Due to this overlap, recent trends have shifted toward a functional definition of EPCs, focusing on their ability to form colonies in vitro and induce angiogenic responses in vivo; this functional characterization helps define EPCs more accurately.
When studying nicotine’s impact on EPCs in vivo, researchers observed increased recruitment and angiogenic activity of EPCs due to systemic nicotine exposure in a mouse hindlimb ischemia (HLI) model [52]. They also used a mouse parabiosis model, connecting a wild-type mouse with a tie2-LacZ transgenic mouse, demonstrating enhanced mobilization and recruitment of EPCs. Nicotine-treated mice exhibited higher EPC numbers and enhanced EPC migration ex vivo. Likewise, Sugimoto et al. reported improved EPC efficacy when delivered intravenously in a mouse HLI model [53]. In their study, mice received nicotine through drinking water one week prior to HLI and throughout a 3-week experiment.
While these studies provide compelling evidence for short-term nicotine exposure improving EPC numbers and function in vivo, long-term exposure studies have shown contrasting results. In one such study, mice received systemic nicotine via drinking water for up to 6 months [54], a notably longer duration than the 3-week exposure In Sugimoto study et. al., study. After one month of nicotine exposure, EPC number and activity resembled earlier short-term exposure findings. However, after 3 and 6 months, EPC numbers significantly decreased, and cell proliferation and migration functions were impaired compared to control EPCs without nicotine treatment. The study also found that telomerase expression and activity decreased after six months of nicotine exposure. These findings suggest that chronic nicotine exposure has detrimental effects on EPC number and function, potentially reducing their therapeutic potential (Table 1).
| Cell Type | In vivo/In vitro | Nicotine Dosage | Exposure Duration | Outcome | Reference |
| Endogenous mice EPCs | In vivo, mouse parabiosis model with HLI model | 100 μg/ml in drinking water or intramuscular injection (0.03 μg/kg) | 30 days |
|
[52] |
| Human EPCs from healthy nonsmokers | In vitro, media | 2 μM | 48 h |
|
[53] |
| Mouse EPCs | Ex vivo | 100 ng/ml in drinking water | 3 weeks |
|
[53] |
| Mouse EPCs | In vivo (mice HLI model) | 100 ng/ml in drinking water | 5 weeks |
|
[53] |
| Human EPCs from umbilical cord blood | In vitro | 10 nM | 36 h |
|
[62] |
| Mouse EPCs | Ex vivo | 100 ng/ml in drinking water | 1, 3 and 6 months |
|
[54] |
| Human MSCs | In vitro | 0.1, 1 and 10 μM | 24 h |
|
[55] |
| Human MSCs | In vivo | 0.01 μM, iv. | 24 h |
|
[55] |
| Rat BM MSCs | In vitro | 25, 50 and 100 μM | 4 weeks |
|
[60] |
| Human MSCs | In vitro | 0.1, 0.5 and 1 μM | 48 h |
|
[58] |
| Human MSCs and periodontal ligament derived stem cells | In vitro | 1 μM | Up to 7 days |
|
[56] |
| Human umbilical cord derived MSCs | In vitro | 0.1, 0.5, 1 and 1.5 mg/ml | 2 h |
|
[57] |
Table 1: Nicotine exposure and its effect on multipotent stem cells.
Mesenchymal stem cells are versatile adult stem cells that were initially isolated from bone marrow, but have since been found in multiple tissues like adipose tissue, dental tissue, skin, umbilical cord, etc. MSCs are extensively studied in clinical stem cell therapies, due to their easy isolation from bone marrow and adipose tissue. Given the numerous clinical trials involving MSCs, considering the impact of lifestyle choices such as smoking, and nicotine exposure is crucial when evaluating their therapeutic effectiveness [33].
In vitro effects of nicotine on MSCs have been extensively explored, with variations in dosage and exposure duration across studies. Schraufstatter, et. al., reported a significant increase in apoptosis in human MSCs after a short 24-hour exposure to 1 μM nicotine, by utilizing propidium iodide staining [55]. This study also revealed inhibited chemotactic migration (Table 1). Similar findings were replicated in a study comparing human MSCs with periodontal ligament-derived stem cells, where both cell types exhibited impaired proliferation and migration as early as one day after nicotine exposure to 1 μM nicotine over 5 to 7 days [56]. Additionally, exposure to 6.2 mM nicotine resulted in increased apoptosis and alterations in cell morphology, including enhanced vacuole formation in the cytoplasm and pyknotic nuclei [57]. Proteomic analysis unveiled differential expression of markers associated with cell structure and movement, which could contribute to the observed impairments in migratory functions following nicotine exposure [57]. This concept was further explored in an investigation into nicotine’s impact on the mechanical properties of MSCs, revealing that cells became significantly stiffer than untreated controls after a 48-hour exposure regimen, indicating reduced motility [58].
Studies conducted by Wahl et al. found that differentiation efficacy was another vital assessment. Such findings showed nicotine alone mirrored results obtained with cigarette smoke; revealing the difference in effects of cigarette smoke and nicotine alone are minimal. Media infused with cigarette smoke decreased proliferation and migration of human adipose-derived MSCs [59]. The authors also found a decrease in the secretion of inflammatory markers IL6 and IL8, crucial for chemotaxis in wound-healing processes. The chondrogenic differentiation of MSCs was also notably hampered in response to nicotine exposure, with the extent of impairment depending on concentration [56,60].
In vivo studies of MSCs exposed to nicotine have demonstrated impaired homing of intravenously delivered MSCs to the bone marrow and spleen in mice, while the lung remained unaffected [55]. Thus, validating other in vitro findings showing decreased migratory function of MSCs post nicotine exposure. In order to assess osteogenic function, Zhao et al. isolated alveolar bone marrow MSCs from healthy nonsmokers and smokers [14]. In a subcutaneous murine model, polylactic acid scaffolds seeded with MSCs were assessed for bone matrix after 8 weeks, revealing that MSCs from smokers generated substantially less bone matrix when compared to those from nonsmokers. Four weeks after autologous scaffold transplantation in patients, higher implant stability was found in nonsmokers. In a direct comparison between adipose-derived MSCs obtained from individuals who smoke and those who do not, Barwinska and colleagues employed a mouse hindlimb ischemia model. Their findings indicated that MSCs derived from smokers displayed a marked reduction in angiogenesis when compared to those from nonsmokers [61,62]. Within the same study, in vitro experiments conducted hinted at a potential molecular mechanism, suggesting that activin A might be involved, leading to decreased secretion of HGF and stromal-derived factor-1 (Table 1).
Embryonic stem cells (ESCs) are highly versatile pluripotent cells derived from the inner cell mass of embryos. They express surface markers like stage-specific embryonic antigen-4, Tra-1-60, Tra-1-81, and pluripotent markers such as Oct3/4. They are known for their potential in disease modeling and preclinical studies, including spinal injuries, cardiovascular disease, and neurological disorders [63,64]. However, ethical concerns surrounding viable embryo destruction have led to a decreased use of human ESCs in regenerative medicine [65].
ESCs serve as valuable tools for studying embryonic development in the context of tobacco and nicotine exposure and research utilizing mouse and human ESCs has demonstrated the detrimental effects of this exposure. By comparing regular and harm-reduction cigarettes, increased cytotoxicity and impaired adhesion in both mouse and human ESCs after 24-hour exposure to mainstream or secondhand smoke was revealed [66,67], with secondhand smoke exhibiting higher cytotoxicity. Additionally, experiments with nicotine alone showed that low concentrations (0.01 and 0.1 μM) promoted mouse ESC proliferation, while high concentrations (1 and 10 μM) hindered proliferation after 48 hours [68]. These studies also revealed dysregulation of the Wnt signaling pathway due to nicotine exposure. Furthermore, in vitro investigations demonstrated impaired cardiac differentiation of human ESCs when exposed to cigarette smoke and e-cigarette smoke, resulting in reduced expression of contractile and junction proteins, ultimately affecting cardiomyocyte maturation [69].
Due to their ability to be generated autologously for regenerative medicine uses, induced pluripotent stem cells (iPSCs) have become increasingly popular when considering the effect of nicotine on iPSC differentiation and proliferation [33]. Similar to the in vitro experiments from the prior studies mentioned of other cell types, the experiments performed by Ishizuka et al. shows that nicotine exposure on a short-term level led to increased proliferation of iPSCs [70]. However, increasing the concertation of e-liquids at concentrations of 0.1% and higher caused human iPSC proliferation to greatly decrease and a greater generation of reactive oxygen species and apoptosis was witnessed [39].
Within the same investigation, endothelial cells derived from iPSCs were subjected to serum from individuals who were nonsmokers, cigarette smokers, and e-cigarette users. E-cigarette usage displayed effects on endothelial dysfunction and the generation of reactive oxygen species that closely resembled the impact of cigarette usage, thus highlighting the noteworthy implications of nicotine exposure in its various manifestations.
In a peripheral arterial disease experiment, mice not exposed to chronic nicotine responded positively to induced pluripotent stem cellderived endothelial cell implantation in their ischemic limbs, leading to improved perfusion recovery. However, mice with chronic nicotine exposure did not respond to this therapy. [72]. Ma, et al., recently established a rabbit model for mandibular lengthening, designed to evaluate the impact of nicotine on bone regeneration. This model employed 60-day time-release nicotine pellets [5]. When compared to alternative nicotine delivery methods, the surgical implantation of the time-release nicotine pellets placed subcutaneously offered a dependable and convenient means to maintain a stable blood nicotine level over an extended period [5].
The influence of nicotine on bone healing in distraction osteogenesis was found to be dose dependent. At lower plasma nicotine levels, no significant adverse effects on bone healing were observed, although histological examinations revealed that the newly formed bony trabeculae were not as mature as those in the control groups. In contrast, exposure to high doses of nicotine significantly compromised bone regeneration in distraction osteogenesis [5]. This model provides a platform for further research into the mechanisms underlying nicotine’s effects on bone healing.
A study on the effects of nicotine exposure on neural stem and progenitor cells presented findings suggesting that e-cigarette (EC) liquids and aerosols can result in notable and potentially irreparable harm to the mitochondria of neural stem cells (NSCs). The decline in mitochondrial stem cells could be the reasoning for accelerated aging that is topically observed in chronic smokers. EC treatments resulted in changes in mitochondrial membrane potential (MMP), triggering the clustering of mitochondrial nucleoids. This caused damage to mitochondrial DNA (mtDNA), while inducing an influx of calcium; ultimately leading to retraction of the cell membrane and an overload of intracellular calcium [73].
Siggins, et al., exploited a mouse model to explore the effects of cigarette smoke exposure on hematopoietic stem and progenitor cells (HSPCs) and the hematopoietic stem cell niche within the bone marrow [74]. The findings indicated that smoke exposure leads to a depletion of HSPCs and long-term repopulating hematopoietic stem cells (LT-HSCs). The reduction in LT-HSCs was made evident by specific marker phenotypes, including Lin-c-kit+Sca-1+ (LKS) and signaling lymphocyte activation molecule (SLAM; Lin-CD48- CD150+). They found that hematologic changes in cigarette smokers resulted from niche cell damage, specifically affecting MSCs and their progeny. Smoke exposure reduced both HSPC and MSC counts, which correlated with diminished naïve HSPC engraftment post-bone marrow transplantation in smoke-exposed animals. Despite these reductions, smoke exposure also disrupted MSC gene expression of pro-proliferative signaling proteins. This data indicated that smoke exposure not only reduces the number of MSCs but also dysregulates hematopoiesis by interfering with MSC signaling pathways. These observations may have clinical implications when considering bone marrow transplants for donors with a smoking history [74, 75].
The ability to undergo self-renewal is a fundamental characteristic of stem cells, crucial for their effectiveness in medical applications. Wahl et. al. conducted experiments to investigate the impact of cigarette smoke extract (CSE), which is composed of all the elements contained in cigarette smoke and not just nicotine, on human adiposederived mesenchymal stem cells (MSCs) in vitro, employing varying concentrations. At levels of 5% and 10% CSE, MSC viability was severely compromised, while at 1%, no significant difference in cell viability was observed. This study underscores that smoking can be highly toxic to MSC populations above specific thresholds [59]. However, it is essential to note that this data provides empirical insights, as the concentrations of specific compounds in the extract remain unknown.
In another in vitro study, human umbilical cord blood cells exposed to nicotine concentrations ranging from 0.5 to 1.5 mg/ml (equivalent to 3-9 mM) exhibited dose-dependent reductions in proliferation and increased apoptosis (p < .05 for all concentrations). Notably, significant decreases in proliferation were observed even at lower concentrations, as low as 0.1-10 µM (p<.05). Diminished proliferation rates were also observed in MSCs obtained from chronic smokers without additional exposure to smoke or nicotine in vitro following isolation. These cells demonstrated a 2.53-fold decrease in proliferation compared to control cells from non-smokers. Importantly, reduced proliferation persisted even after sub-culturing the cells 3-5 times, suggesting that the effects of nicotine exposure may be long-lasting or extend through several generations of descendant cells [28]. An evaluation to determine if human clinical trials have been performed investigating nicotine exposure’s effect on stem cell therapy has not been reported to date.
The effect of nicotine exposure on stem cells has been investigated and has shown that different stem cells are affected in various ways. Some cell functions are benefited by the exposure of cigarette smoke and nicotine if it is only for a brief period but if the exposure happens to be chronic or dose-dependent, detrimental effects occur such as decreased proliferation, efficacy of differentiation, migration, and increased apoptosis. Studies such as Wahl, et al., indicated that the outcomes of nicotine alone closely resembled those derived from cigarette smoke, suggesting minimal distinctions in the effects of cigarette smoke compared to nicotine alone [59]. Screening for stem cell donors should be more thoroughly investigated as implicated by observations made when examining bone marrow transplants and donors with a smoking history [58]. The reasoning for the inclusion of the vaping epidemic among adolescents is due to this substance’s detrimental effects on stem cell therapies especially when high doses or chronic use of nicotine are involved. The surge of nicotine addiction among younger generations is not only a pressing public health concern in itself but it also has potential far-reaching consequences. These repercussions involve placing a substantial burden on healthcare professionals specializing in health issues arising from vaping and potentially compromising the feasibility of employing stem cell therapies to reverse the damage adolescents have done to their health. This is particularly concerning due to extensive potential of stem cell therapies in the medical field. Therefore, addressing the surge in nicotine addiction among adolescents becomes not only a public health imperative but also a crucial step in preserving the potential of stem cell therapies for the future.
Download Provisional PDF Here
Article Type: REVIEW ARTICLE
Citation: Karges A, Inayat MS, Gallicchio VS (2024) Stem Cells and Nicotine Exposure: A Growing Concern for Adolescent Health. J Clin Lab Med 7(1): dx.doi.org/10.16966/2572-9578.143
Copyright: © 2024 Karges A, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access