Introduction
Caries is a multifactorial disease [1] where bacterial acid formation from sugars and pH-neutralising alkali formation are important factors by regulating the dental biofilm ecosystem [2]. An imbalanced ecosystem with an acidification exceeding the alkalisation may result in a demineralisation of the tooth surface [3]. However, the balance could be restored through the salivary buffering system and alkali (ammonia) production from urea and arginine by oral microorganisms [4].
A three-fold higher urease level [5] and a higher urease activity [3] in caries-free compared with caries-active individuals has been reported and several studies showed higher supragingival plaque-pH in caries-free individuals [5-7]. It was further acknowledged that a higher caries risk is associated with a low alkali-generating capability of oral bacteria [3].
Several in vitro studies have shown that the dental plaque bacteria can adapt to low pH values from frequent exposure to sugars [8]. The in vivo adaptation of plaque bacteria to an acidic and alkaline environment was also reported [6,9,10]. There is little published information about bacterial adaptation to alkaline and their capacity to adapt to acid and alkaline environment if the bacteria are isolated from caries-active and caries-free individuals respectively.
In a study by Hassan H, et al. [11] the supragingival plaque pH from sugar fermentation in vivo in caries-free and caries-active individuals was examined after the adaptation of the plaque microbiota to acid and also to alkali by one-week frequent rinsing with sugar and urea respectively. Adaptation to acid had the most pronounced effect on plaque-pH in those with no caries (caries-free). Adaptation to alkali resulted in slightly increased plaque-pH values in those with caries (active caries).
From that study it was hypothesised that the final pH from acid formation by plaque bacteria after acid and alkali adaptation would differ if the bacteria were isolated from caries-free and caries-active individuals respectively. The aim of this pilot study was to assess (in vitro) the effect of adaptation of acid-tolerant supragingival plaque bacteria from caries-free and caries-active individuals on the final pH after sugar fermentation.
Material and Methods
Bacterial strains
The study included a total of 128 bacterial isolates previously obtained after culturing of supragingival interproximal plaque samples on pH 5.2 agar plates [11]. The plaque samples were collected from 10 Caries-free (CF) and 10 Caries-Active (CA) young individuals (15-21 years), before and after one-week adaptation periods with daily rinses 5 times/day with 10% sucrose and 0.25% urea solutions respectively. For isolation, colonies representing different morphology were separately collected and cultured on blood agar for purity control before kept on beads at -80°C. Isolates from beads were cultured on blood agar (Blood Agar Base No.2 CM 271; Oxoid, Basingstoke, UK with 5% defibrinated horse blood and 0,5% Na-lactate) in an atmosphere of 90% N2 and 10% CO2 at 36°C and identified according to gram-staining and characteristic colony morphology after growth on selective Mitis-Salivarius (MS) agar (Acumedia, Neogen Corp., USA), Mitis Salivarius-Bacitracin (MSB) agar [12], and Rogosa SL (RSL) agar (Acumedia) for identification of various streptococci and lactobacilli, respectively. The species identification of the streptococcal isolates was finally confirmed using the API system (API 20 Strep, Biomerieux, Marcy l´Etoile, France).
After culturing overnight on blood agar, bacterial colonies were inoculated and grown in Brain Heart Infusion broth (BHI, Acumedia) for the fermentation test. The bacteria were harvested during logphase, centrifuged at 3500 rpm for 5 minutes and washed twice with Phosphate-buffered Saline (PBS) to eliminate the culture medium. The washed bacteria were diluted with PBS to reach an Optical Density (OD) of 1 (650 nm) as measured in a spectrophotometer, which corresponds to 109 cells/ml.
Test solutions and pH indicator
The sugars used in the fermentation tests were sterile filtered aqueous solutions of 2% sucrose (Difco, Becton, Dickinson and Company, Sparks, MD, USA), glucose (Difco), fructose (Merck, Darmstadt, Germany) and lactose (Merck). The pH indicator used, bromocresol purple (Svenska Finkemikalier, Sweden), was prepared in a 0.3% aqueous solution which turns yellow at pH <5.2 and purple at pH >6.8 according to the manufacturer.
Fermentation assay
Microtiter plates (96 MicroWell™ Plates Nunclon™; Nunc, Roskilde, Denmark) were used in the tests, which were performed as previously describe [13,14]. All sugars were tested in duplicate. In each well 50 μl of BHI broth, 50 μl of sugar, 10 μl of the bacterial solution and 10 µl of indicator solution were added. Wells with PBS instead of bacterial suspension was used as control. The plates were covered with laboratory plastic foil and incubated in 36°C in an atmosphere of 10% CO2+90% N2. Photos for registration of the colour after 24 hours of incubation were taken and saved to document the colour changes of all tests at one and the same occasion. The tests were conducted and individually evaluated by two examiners who were unaware of isolate type, the source of isolates (CA or CF) or the test period (before/after acid/alkali adaptation). Wells representing different colours were also tested using pH sticks and it was concluded that colours representing the pH-intervals <5 (yellow), 5-6 (colour in between yellow to purple) and >6 (purple) could clearly be discerned.
Statistical analyses
Significance of differences between the groups regarding bacteria and within the groups regarding acid formation from sugars before and after adaptation were analysed by using SAS 9.1.3 (SAS Institute Inc, NC, USA) and Pearson’s Chi2-test.
Results
Altogether, 68 isolates of Streptococcus mitis, 37 Streptococcus oralis, 4 Streptococcus mutans, 3 Streptococcus sanguinis, 3 Streptococcus salivarius and 13 Lactobacillus spp. were identified. The non-mutans streptococci e.g. S. mitis and S. oralis dominated among the acidophilic isolates in both CA and CF individuals and constituted 53% and 29% respectively of total 128 isolates (Table 1). It should be noted that the only occasion where these streptococci did not dominate was in the CA group (4 out of 14 isolates) after acid adaptation (AAA). Furthermore, Lactobacillus spp. was found in this group only, both before (BAA/ BBA) and after (AAA/ABA) the respective adaptation periods, and other bacteria, S. mutans, S sanguinis, and S. salivarius, were isolated more sporadically. This finding of bacteria was significantly different between CF and CA individuals (p<0.05, before and p<0.01 after acid adaptation).
|
Bacterial Species |
CF |
CA |
CF |
CA |
| BAA* (n=20) |
AAA# (n=20) |
BAA* (n=20) |
AAA#
(n=14) |
BBA*
(n=19) |
ABA
(n=14) |
BBA*
(n=10) |
ABA
(n=11) |
| Streptococcusmitis |
10 |
8 |
10 |
4 |
10 |
11 |
7 |
8 |
| Streptococcusoralis |
10 |
12 |
3 |
0 |
9 |
3 |
0 |
0 |
| Streptococcusmutans |
0 |
0 |
2 |
1 |
0 |
0 |
1 |
0 |
| Streptococcussanguinis |
0 |
0 |
1 |
2 |
0 |
0 |
0 |
0 |
| Streptococcussalivarius |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
0 |
| Lactobacillus spp. |
0 |
0 |
4 |
4 |
0 |
0 |
2 |
3 |
Table 1: Bacterial isolates obtained from caries-free (CF) and caries-active (CA) individuals, before (BAA) and after (AAA) acid adaptation, and before (BBA) and after (ABA) alkali adaptation respectively.
*Indicates statistical significant difference (p<0.05) between type of isolated bacterial species from CF vs. CA individuals before acid and base adaptation (Chi2 test).
#Indicates statistical significant difference (p<0.01) between type of isolated bacterial species from CF vs. CA individuals after acid adaptation (Chi2 test).
Figure 1 shows the proportions of isolates (%) that could form acid to pH <5 or between pH 5 and 6, before (BAA) and after (AAA) acid adaptation (Figures 1A and 1B) with sucrose, glucose, fructose and lactose in the CF and CA individuals respectively. The pattern was consistent for all sugars in the way that the proportion of isolates forming acid to pH <5 increased in both groups (CF and CA) and to highest extent in the CF group, however not enough to reach statistical significance for each of the sugars. Before both adaptation periods the proportion was higher in the CA group than in the CF group. After adaptation to alkali, a light decrease in % isolates at pH <5 was seen for sucrose and glucose in the CF group (Figure 1C) and more consistently and clearer in the CA group (Figure 1D), although not significant in the two groups for any of the sugars. Statistical analysis was not performed between the groups due to use of non-comparable units (four different sugars).
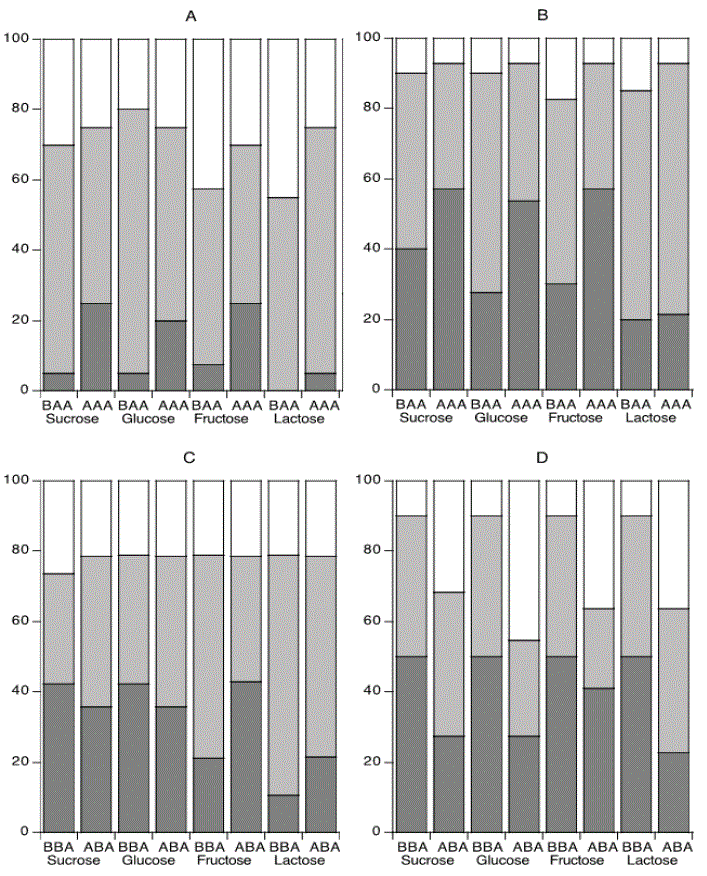
Figure 1: Distribution of bacterial isolates (%) between the different pH intervals <5, 5-6 and >6 after fermentation of sugars.
The bacteria were isolated before (BAA) and after acid adaptation (AAA) from the caries-free (Figure A) and caries-active (Figure B) individuals, and before (BBA) and after alkali adaptation (ABA) from the caries-free (Figure C) and caries-active (Figure D) individuals. Statistical analysis could not be performed between the groups due to use of non-comparable units (four different sugars).
Discussion
The present single-blinded in vitro study assessed the effect of adaptation to acid and alkali on the ability of acid tolerant plaque bacteria to lower pH after sugar fermentation. The bacteria were isolated after culturing plaque samples from caries-free (CF) and caries-active (CA) young individuals on pH 5 agar [11].
Both before and after adaptation to acid and alkali, the CF individuals were characterised by non-mutans streptococci Streptococcus mitis and Streptococcus oralis, previously reported to be frequently found in the caries-free biofilm ecology with a controlled acidic environment [15,16]. The CA group was characterised by more of acidogenic and aciduric lactobacilli and mutans streptococci that thrive in acidic environments [17,18].
The finding of an increased proportion of isolates forming acid to pH <5 after acid adaptation in especially the CF group suggest that the non-mutans streptococci had adapted to the new environment and become more acidogenic. This is in line with the ecological plaque hypothesis [16,17,19] implying that frequent access of sugar may initiate a shift of the biofilm ecology favouring more acidogenic and aciduric phenotypes of bacteria.
Acid adaptation had less effect on the CA group with a lower increased proportion of isolates forming acid to pH <5. Instead, reduced proportions of isolates forming acid to pH <5 after alkali adaptation was most clearly seen among the CA isolates. These findings may be explained by a higher acid tolerance together with a higher ability to metabolise urea and produce ammonia, as of commencement in the CA bacteria as compared with the CF bacteria. Thus, the CA isolates could be expected to be more prone to form acid and alkali than are the CF isolates already prior to the adaptation periods. This complies with a higher proportion of isolates forming acid from the sugars in the CA group before both adaptation periods (Figure 1). The overall pattern was consistent for all experimental groups and all sugars (Figures 1A-1D) although statistical significance was not reached for each of the sugars.
The number of individuals included in the initial study [11] from which the bacteria were obtained was too low to reach statistically significant differences between the groups for each of the sugars tested, which is a limitation of the present study.
The outcome of this in vitro study validated the findings of the in vivo changes of interproximal supragingival plaque-pH before and after acid adaptation. The Stephan curve displayed a greater decrease of the pH-level in the CF group in comparison to the CA group [11]. On the contrary, increased pH-values were seen post-alkali adaptation for the CA group in vivo, corroborating a decreased proportion of bacteria forming acid that could lower pH to <5 in vitro.
Conclusion
In conclusion, the findings suggest that the bacterial ability to ferment sugars resulting in a low environmental pH (<5) to highest extent increases in caries-free individuals after acid adaptation and decreases in caries-active individuals after alkali adaptation.
Competing Interests
There are no potential conflicts of interest for any of the authors. The authors alone are responsible for the content and writing of this paper.
Ethical Approval
The previous study in which the bacterial isolates were obtained [11] was approved by the Ethics Committee at the University of Gothenburg, Gothenburg, Sweden (Dnr 282-10).
Acknowledgement
We thank Miss Fanny Bjondahl and Mr. Richard Olofsson for performing the fermentation procedures and Mrs. Susanne Blomqvist for excellent technical assistance and guidance in the microbiological handling and analysis.
Article Information
Article Type: RESEARCH ARTICLE
Citation: Hassan H, Dahlén G, Carlén A (2020) Final pH from Sugar Fermentation by Plaque Bacteria Isolated from Caries-free and Cariesactive Individuals after Frequent Sugar and Urea Rinses. Int J Dent Oral Health 6(3): dx.doi.org/10.16966/2378-7090.319
Copyright: © 2020 Hassan H, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
Received date: 12 Dec, 2020
Accepted date: 05 Mar, 2020
Published date: 12 Mar, 2020
